

1. 서 론
2. 재료 및 방법
2.1 정점 설정
2.2 대형저서동물 채집
2.3 실험실 분석
2.4 환경요소 분석
3. 결과
3.1 환경 특성
3.2 종 조성
3.3 종수, 서식밀도, 생체량의 시공간 분포
3.4 주요 우점종
3.5 집괴분석
3.6 환경요소와의 관계
4. 고 찰
4.1 공간분포 특징
4.2 주요 우점종의 조성 및 분포
4.3 군집 구조
5. 결 론
1. 서 론
하구역은 바다와 강이나 하천이 만나 자유롭게 담수와 해수가 혼합되는 장소로서 다양한 생태학적 과정이 일어나며, 해양생물들의 이동통로로서 기수역에는 고유한 생물이 서식하는 생태계의 중요한 장소이다(Copland and Bechtel, 1974; McLusky and Elliott, 2004). 이곳은 담수 유입에 의한 염분 저하 및 퇴적상의 변화 등이 가장 큰 환경 요소로서 작용하기 때문에 기본적으로 염분 및 퇴적상에 의해 저서동물 군집이 조절되는 장소이다(Boesch et al., 1976). 또한 조석 작용은 염분 변화를 극대화하여 정착성 저서동물에게 큰 영향을 미친다(Ritter et al., 2005). 하구역에서 담수의 자유로운 유입과 교환이 차단될 경우 하구역 고유의 성질을 상실하게 되고 그 영향은 하구역 생태계 전반으로 미칠 수 있다. 대부분의 하구역에는 항구가 발달하고 인구 밀집으로 인한 유기오염 문제가 대두되어 하구역 고유의 특성이 상실된 곳이 많이 존재한다(McLusky and Elliott, 2004).
우리나라에는 중요한 5개 하천(한강, 금강, 영산강, 섬진강, 낙동강)에 하구역이 발달해 있으며 낙동강, 영산강, 금강은 바다에 인접하여 하구둑을 건설하여 주변 육지를 농경지 및 공업단지로 사용하고 담수와 해수의 자유로운 혼합을 차단하여 하구역 기능이 훼손된 곳이다. 하구둑이 없어 담수와 해수가 혼합되는 자연성을 띤 강은 한강 및 섬진강이 있으며, 그 밖에 바다로 직접 연결되어 있는 소하천들이 있다. 이러한 장소는 해수가 상류까지 침투할 수 있고 염분 및 퇴적상 변화에 따라 상류에서 하류에 이르기까지 특징적인 생물 분포를 나타낸다(Lim et al., 2012). 한강 하구역의 경우 접경지역이기 때문에 영종도 주변을 중심으로 정점을 설정하고 현장 조사를 수행한 경우가 대부분이기 때문에 하구역의 뚜렷한 생물 분포 양상을 파악하기 어려웠다(Hong and Yoo, 1996; Lim et al, 1995; Shin et al., 1989, 1992). 그러나 금강 하구역의 경우 Choi and Koh(1994)가 새만금 방조제가 건설되기 이전의 상태에서 저서동물 군집을 조사하고 일차적으로는 염분 농도, 이차적으로는 퇴적상에 의해 군집이 조절됨을 밝혔으며 특징적인 생물 분포 양상을 파악하였다. 낙동강 하구역의 경우 Lee et al.(2005)이 2001년 9월의 1회 조사를 통해 대형저서동물 군집은 퇴적상과 유기탄소량 및 수심에 따라 영향을 받는 것으로 보고하였다. 따라서 하구역의 특징인 염분 및 퇴적상의 차이에 따라 뚜렷한 시공간분포 양상을 보이는 연구는 주로 소형 하천에서 수행된 결과들이다. 즉, 남대천 하구역(Hong et al., 2004), 법성포 와탄천 하구역(Lim and Hong, 2002), 수어천 하구역(Lim et al., 2012) 등에서는 염분 농도 경사와 퇴적상에 따른 저서동물 군집의 공간 분포에 대한 상세한 연구가 수행되었다.
하구역 생태계를 안정적으로 관리하기 위해서는 이 해역의 생태계 요소에 대한 시공간 분포 특성을 지속적으로 파악하여야만 하고, 이를 바탕으로 관리 계획을 수립하여야 한다. 그러나 한강, 금강, 낙동강 하구역에서 수행된 결과들은 모두 특정 계절에 국한된 일시적인 조사이거나 1년간에 걸친 계절 조사였기 때문에 이 해역에서의 장기적인 환경 변화를 모니터링 할 수 있는 자료를 획득하는 데는 한계가 있었다. 또한 낙동강 하구역에 포함되는 마산만의 경우 Lim and Hong(1997)이 3년간에 걸친 진해만 저서동물 군집 조사를 통해 군집 구조, 빈산소 수괴 형성, 우점종의 공간분포 등을 파악하였으나 조사장소는 진해만에 주로 국한되었으며 낙동강 하구역까지는 포함하지 못하였다.
본 연구에서는 우리나라 주요 하천인 한강, 금강, 낙동강 하구역에 대한 해양생태계 관리 계획 수립을 위한 기초 조사의 일환으로 저서동물의 군집구조와 주요 우점종의 시공간 변동 양상을 파악하였다. 또한 저서동물 군집 구조의 결정에 미치는 주된 환경 요소를 파악하여 각 하구역 저서생태계의 향후 중점 관리방안을 제안하였다.
2. 재료 및 방법
2.1 정점 설정
국가 해양생태계 종합조사의 일환으로 우리나라의 주요 하천인 한강, 금강, 낙동강 하구역에서 대형저서동물 채집을 위해 총 20개의 정점을 설정하였다. 한강 하구역에서는 영종도 북부에서부터 자월도 및 영흥도 북부해역까지 7개 정점, 금강 하구역에서는 새만금 방조제 외측까지 6개 정점, 낙동강 하구역에는 가덕도 외측과 진해 마산만을 포함하여 7개 정점을 설정하였다. 각 정점은 강으로부터 유입되는 담수의 영향을 파악할 수 있도록 조사선 형태로 설정되었으며, 주변에 인접한 시화방조제, 새만금방조제 및 마산만의 환경 특성을 고려하여 배치되었다(Fig. 1).
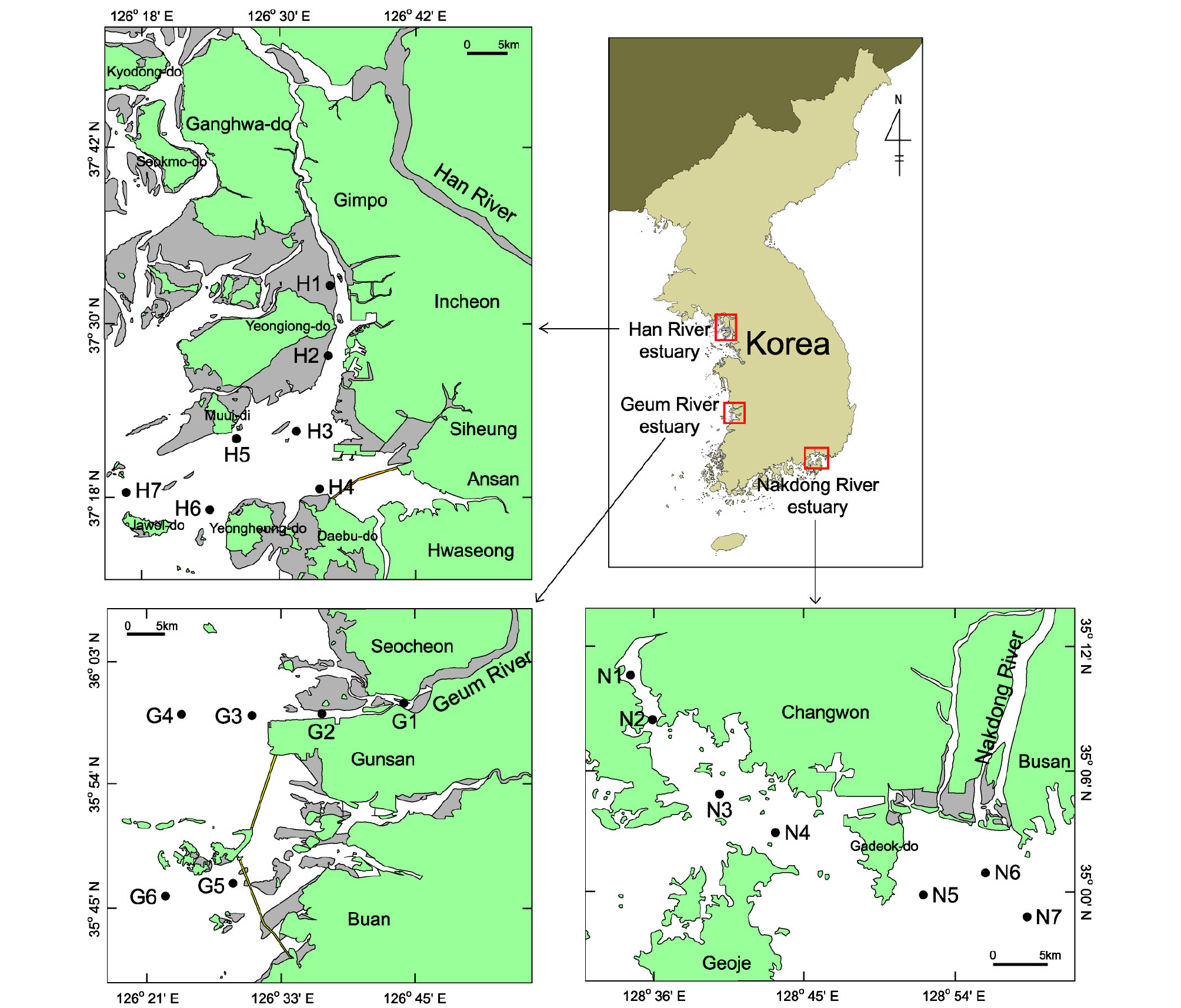
2.2 대형저서동물 채집
현장조사는 2015년, 2016년 및 2017년의 겨울철(2월)과 봄철(5월∼6월), 여름철(7월∼9월), 가을철(11월)에 걸쳐 총 12회가 수행되었으며 조사된 총 정점수는 240개이다(Table 1). 대형저서동물 채집은 van Veen grab (채집면적 0.1 m2)을 사용하여 각 정점당 3회씩 반복 채집하였다. 채취된 퇴적물은 선상에서 1 mm 표준체에서 체질하여 펄을 제거하였고, 잔존물을 시료병에 담고 10% 중성포르말린 수용액으로 고정한 다음 실험실로 운반하였다.
Table 1. Sampling month of macrozoobentos in the study area, from February 2015 to November 2017
2.3 실험실 분석
운반된 퇴적물 시료는 담수로 세척하여 포르말린을 제거한 다음 저서동물을 선별하였으며, 각 분류군별 습중량을 측정하고 종 수준까지 동정하였다. 종 수준까지 동정이 어려운 분류군은 상위분류군까지 동정하였으며, 동정된 종은 개체수를 세고, 단위 면적당(1 m2) 개체수로 환산하여 분석에 사용하였다. 저서동물 군집해석을 위하여 종 다양도(Shannon and Wienner, 1963)를 구하였으며, 집괴분석을 통해 정점군을 구분한 다음 군집을 해석하였다. 집괴분석은 각 정점별 출현종의 개체수 자료를 사용하여 Bray and Curtis(1957) 유사도지수를 구한 다음, 가중평균결합법을 사용하여 수지도를 작성하였다. 또한 집괴분석 결과를 비교 검증하고, 분석의 정확성을 기하기 위하여 다차원척도법(nMDS)을 실시하였다. 구분된 각 정점군의 퇴적환경 및 염분 그리고 우점종을 바탕으로 각 하구역의 대형저서동물 군집 특성을 파악하였다. 또한 각 정점군 구분에 기여한 우점종을 파악하기 위하여 SIMPER 분석을 실시하였다. 저서군집에 미치는 환경요소의 조합을 파악하기 위하여 각 환경자료에 대해 변수값에서 평균값을 뺀 다음 표준편차로 나누어서 표준화(nomalization)한 후 Bio-Env 분석을 실시하였다. 분석된 환경 요소와 생물 요소와의 상관관계는 비모수통계인 Spearman correlation coefficient를 구하여 파악하였다(Primer, v. 6; SPSS, v. 23).
2.4 환경요소 분석
퇴적환경요인으로는 동일 정점에서 동시에 채집된 퇴적물 시료로부터 퇴적물 입도 및 조성비, 강열감량(Loss of Ignition, LOI)을 측정하였고, 수층 환경요인으로는 저층 수온과 염분, 용존산소를 이용하였다. 분석방법은 해양환경공정시험기준(MOF, 2013)을 따랐다. 모든 환경 및 생태자료는 해양환경관리공단 바다생태정보나라와 국가해양생태계 종합조사 보고서에서 취득하였다(KOEM, 2018).
3. 결과
3.1 환경 특성
한강, 금강 및 낙동강 하구역 조사정점에서의 염분 및 평균 입도의 공간 변화는 Fig. 2와 같다. 한강 하구역의 경우 29.3~31.5 psu 범위로서 내측의 정점 H1에서는 상대적으로 낮고 외해역으로 가면서 다소 증가하지만(정점 H4, H5) 큰 차이는 없었다. 그러나 금강 하구역의 경우에는 정점 G1에서 상대적으로 가장 낮은 염분농도를 보였으며 외해역으로 가면서 염분이 증가하는 양상을 보여(정점 G4) 금강 하구에 가까운 정점 G1은 담수의 영향을 상대적으로 크게 받고 있는 것으로 나타났다. 낙동강 하구역의 경우 한강 하구역이나 금강 하구역에 비해 염분이 상대적으로 높고 마산만 내측(정점 N1)에서 외해역으로 가면서 약간 증가하는 양상을 보이지만(정점 N4) 정점간의 차이는 거의 없었다.
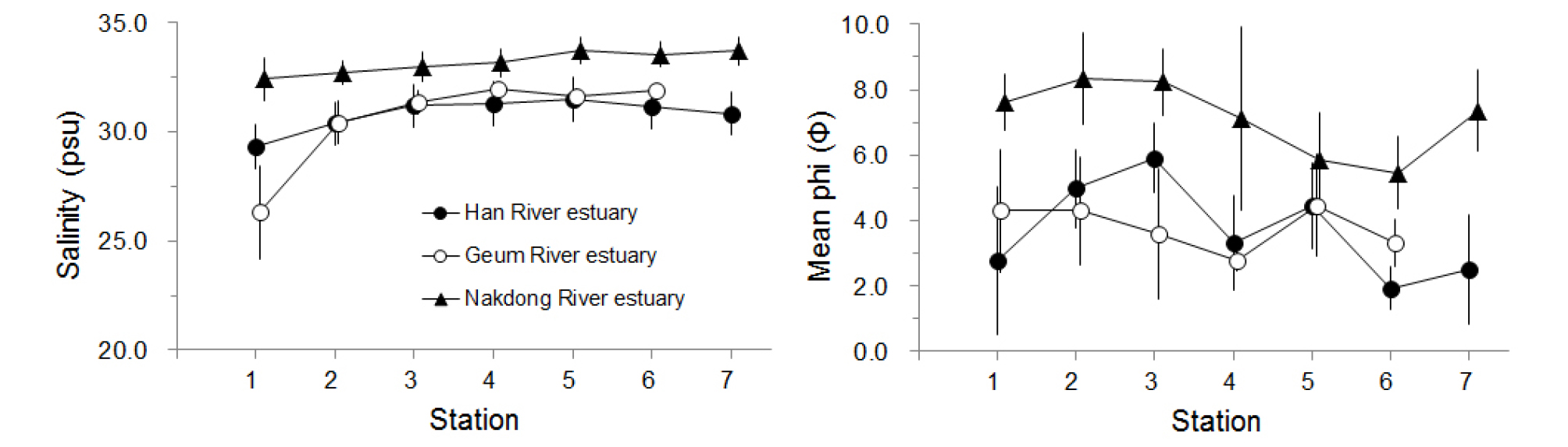
표층퇴적물의 평균 입도는 한강 하구역의 경우 수로역에 위치한 정점 H1에서는 평균 2.8φ로 조립하였으나 인천항 앞에 위치한 정점 H3까지는 입도가 점차 세립해지는 양상을 보였다. 가장 외해역에 위치한 정점 H6과 H7에서는 1.9~2.5φ 범위로서 상대적으로 가장 조립한 양상을 보였으며 모래 함량도 72.8~95.0%로 가장 높았다. 금강 하구역의 경우 금강 입구역의 정점 G1에서는 상대적으로 세립하였으나 외해역으로 가면서 입도가 조립해져 정점 G4에서는 2.8φ로 조립한 양상이었으며 모래 함량은 90% 이상이었다. 낙동강 하구역의 경우 한강 하구역 및 금강 하구역에 비해 세립질 퇴적상을 보였다. 마산만 내측에는 7.1φ 이상의 세립질 퇴적상이었으나 외해역에 면한 정점 N5, N6에서는 5.5~5.9φ로 상대적으로 조립한 양상을 보였다.
3.2 종 조성
조사결과 3개 하구역에서 총 1,008종의 저서동물이 출현하였다(Table 2). 한강 하구역에서 602종, 금강 하구역에서 612종, 낙동강 하구역에서 619종이 출현하여 3개 하구역의 출현종수는 유사한 양상을 보였다. 환형동물의 출현종수 점유율은 3개 하구에서 43~47%를 점유하여 가장 우점한 분류군이었으며, 절지동물은 23~27% 범위의 점유율을 보였다. 연체동물은 한강 하구역에서는 17%의 점유율이었으나 금강 하구역에서는 21%였다. 3개 하구역의 평균 밀도는 1,357 ind./m2였는데 한강 하구역에서 1,127 ind./m2로서 가장 낮았으며 낙동강 하구역에서 1,587 ind./m2으로 가장 높았다. 환형동물의 서식밀도 점유율은 3개 하구역에서 63~83% 범위(평균 74%)를 보였으며 한강 하구역이 가장 높고 금강 하구역이 가장 낮았다. 연체동물과 절지동물 서식밀도 점유율은 6~16% 범위로 낮았다. 3개 하구역의 평균 생체량은 116.8 g/m2이었으며 한강 하구역에서 49.0 g/m2, 금강 하구역에서 129.0 g/m2 및 낙동강 하구역에서 174.2 g/m2이 출현하였다. 한강과 금강, 낙동강 하구역 모두 환형동물이 출현종수와 서식밀도에서 우점 분류군이었다. 또한 한강과 금강 하구역에서는 연체동물이, 낙동강 하구역에서는 극피동물이 생체량 우점 분류군이었다. 종 다양도(H’)는 3개 하구역에서 평균 3.1±0.9였으며 한강 하구역에서 3.1±0.7, 금강 하구역에서 3.2±0.9 및 낙동강 하구역에서 3.2±1.2로서 유사한 값을 보였다.
Table 2. The taxonomic composition of number of species, density, biomass and species diversity (H') in macrozoobenthic community of three major estuaries in Korea, data pooled from February 2015 to November 2017
3.3 종수, 서식밀도, 생체량의 시공간 분포
3.3.1 한강 하구역
출현종수는 겨울철에는 상대적으로 적고 봄철에 증가하여 여름철에 최대값을 보인 후 가을철에 다시 감소하는 매년 반복적인 양상을 보였다(Fig. 3). 2015년에는 각 조사 계절 모두 다른 해에 비해 많은 출현종수를 보였으며 여름철인 8월에는 218종이 출현하여 가장 많은 종수를 보였다. 특히 영종도와 인천 사이의 수로역(정점 H1, H2)과 무의도 남측 정점(H3)에서 많은 출현종수를 보였다. 영종도 주변 해역으로부터 자월도 북측의 외해역으로 나오면서 출현종수가 감소하는 양상을 보인다(Fig. 4).
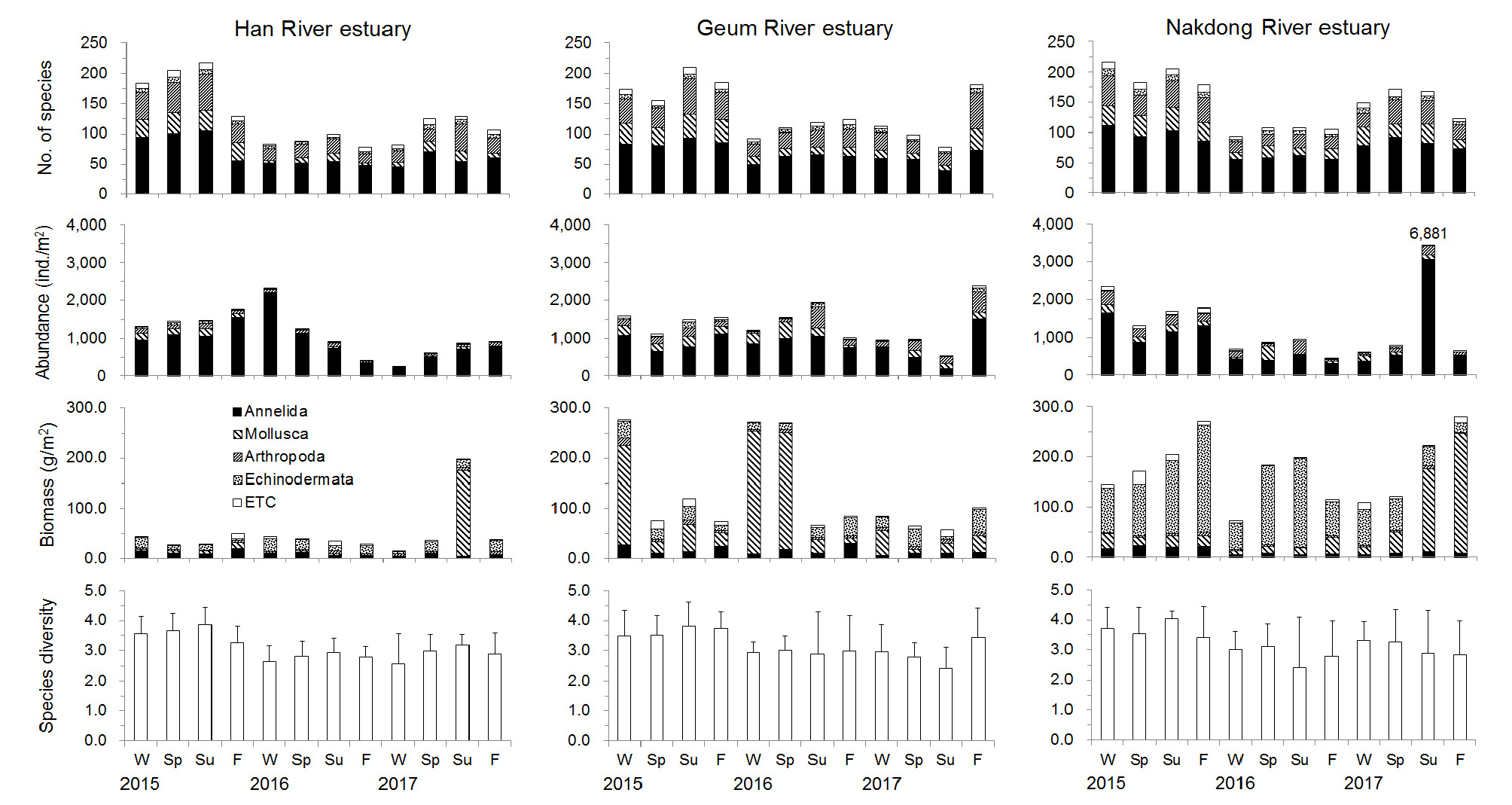
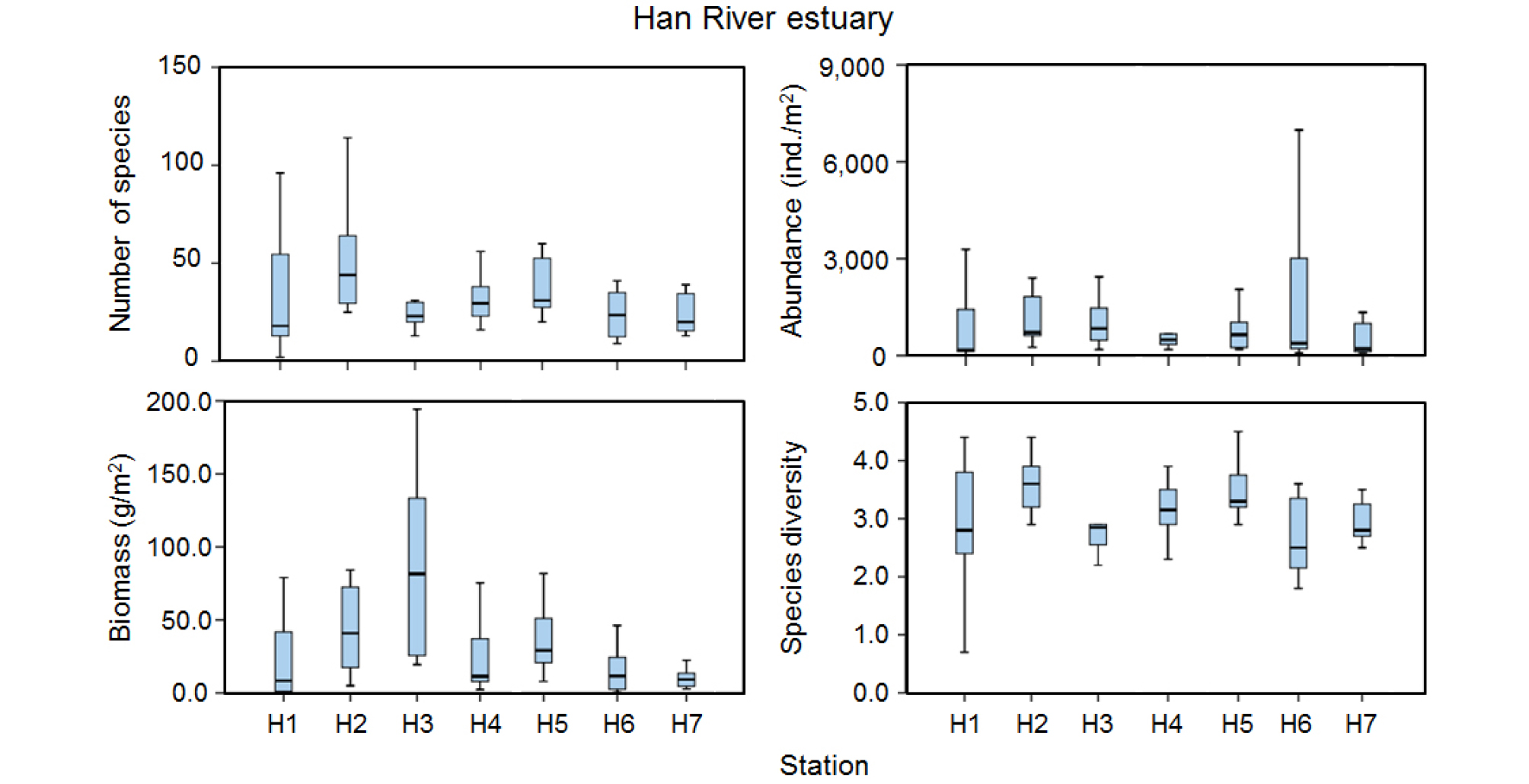
밀도의 경우 2015년 겨울에서부터 2016년 겨울철까지 지속적으로 증가하였으나 봄철인 2016년 5월부터 감소하기 시작하여 겨울철인 2017년 2월에 평균 224 ind./m2로서 가장 낮은 밀도를 보였다(Fig. 3). 공간분포 양상은 자월도 및 영흥도 북측 정점(H6)에서 높은 밀도를 보였는데, 특히 2015년 11월에는 정점 H6에서 전 조사기간 중 가장 높은 9,510 ind./m2의 밀도를 보였다(Fig. 4). 이 정점에서는 다모류 밀도가 96%를 점유하였으며 Aonides oxycephala의 밀도가 이 정점에서 출현한 전체 밀도의 92%를 점유하였다. 출현종수의 공간분포 양상과는 달리 영종도와 인천 사이의 수로역에서는 상대적으로 낮은 밀도를 보였다.
생체량은 여름철인 2017년 8월에 상대적으로 가장 높은 198.4 g/m2이었으며 다른 계절에는 약 51.0 g/m2 미만의 값을 보였다(Fig. 3). 특히 2017년 8월에는 연체동물의 생체량이 86%를 점유하여 가장 높았는데 영흥도 북측의 정점 H4에서 높은 생체량을 나타내었다. 특히 이 정점에서는 이매패류인 우줄기(Barnea davidi) 치패가 223 ind./m2 출현하여 1,174.2 g/m2의 생체량을 보였다. 대부분의 정점들에서 100 g/m2 미만의 생체량을 보였으며, 영종도 남측의 정점 H3과 영흥도 북측의 정점 H6, 수로역의 정점 H2에서는 계절에 따라 200 g/m2 미만의 생체량을 보였다. 영종도 북측으로부터 외측으로 나오면서 뚜렷한 공간분포 양상은 보이지 않았다(Fig. 4).
종 다양도는 출현종수의 계절변동과 유사하게 겨울철에 증가하여 여름철에 최대값을 보인 후 가을철에 감소하는 양상을 보였다(Fig. 3). 또한 담수의 영향을 가장 강하게 받는 정점 H1에서는 종 다양도의 계절 변동이 가장 크게 나타났으며 남측의 정점 H3과 H7에서는 상대적으로 적은 변동을 보였다(Fig. 4).
3.3.2 금강 하구역
금강 하구역에서는 2015년 여름철에 210종이 출현하여 조사기간 중 가장 많았으나 2017년 여름철에는 76종이 출현하여 가장 적었다(Fig. 3). 2016년에는 겨울철부터 가을철까지 지속적으로 출현종수가 증가하는 양상을 보였으나 2017년에는 겨울철부터 여름철까지 지속적으로 감소하다가 가을철에 증가하는 다소 상반된 양상을 보였다. 담수의 영향을 가장 많이 받는 정점 G1에서 가장 적은 출현종수를 보였으며 외해역으로 가면서 점차 증가하여 정점 G3과 G4에서 가장 많은 종수를 보였다(Fig. 5). 특히 2015년 봄, 여름, 가을철에는 이러한 출현종수 차이가 가장 뚜렷하게 나타났으며 다른 계절에는 정점 G3과 G4에서 유사한 양상을 보였다. 고군산군도 남측과 비안도 사이에 위치한 정점 G5와 G6에서는 유사하거나 새만금방조제에 인접한 정점 G5에서 많은 출현종수를 보였다.
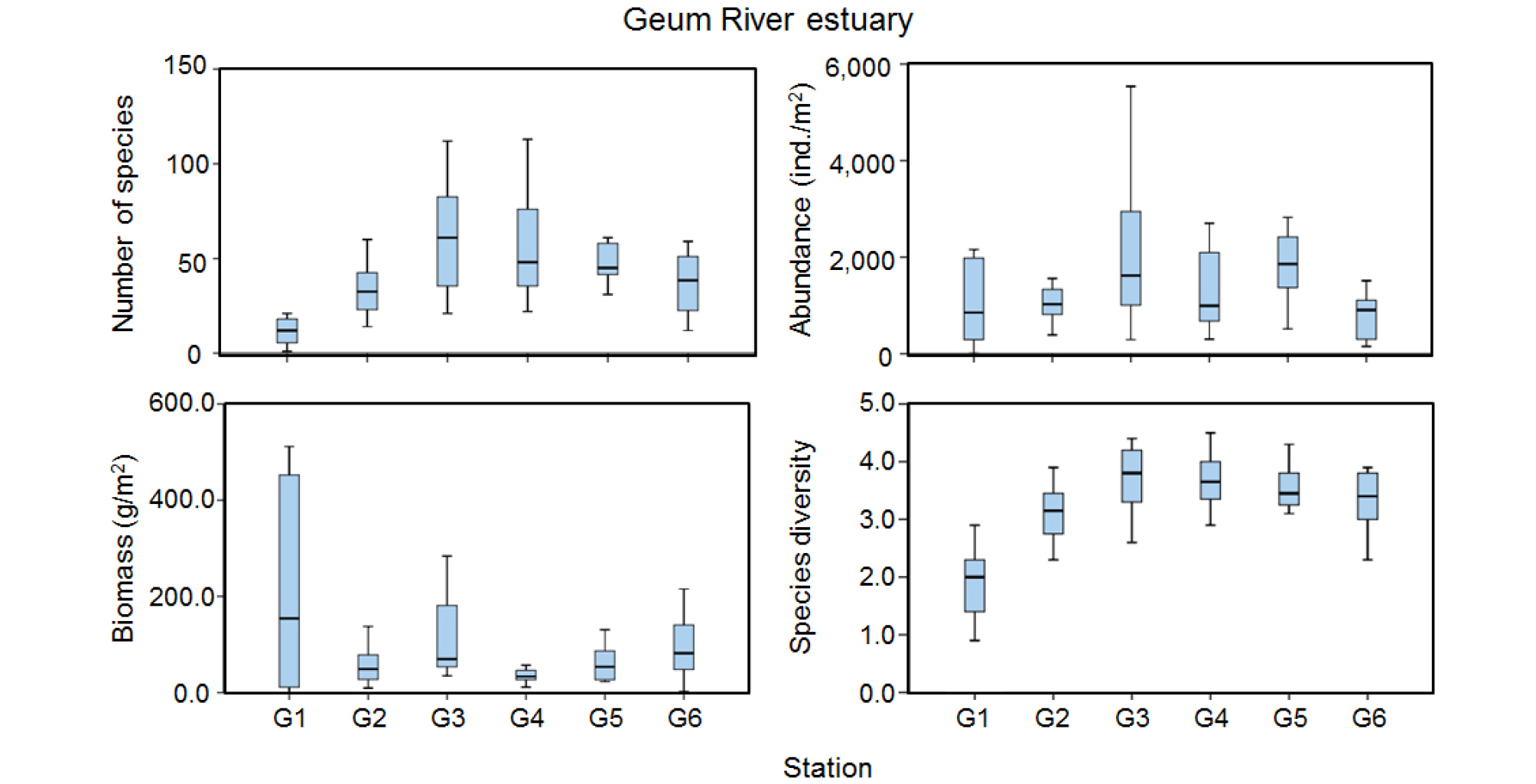
밀도는 계절에 따라 529~2,379 ind./m2 범위였으며, 2016년 겨울철부터 여름철까지 지속적으로 증가하다가 가을철에 감소하였다. 2017년에는 여름철에 가장 낮은 밀도였으며 가을철에 가장 높아 출현종수 계절변동과 유사하였다(Fig. 3). 공간분포 양상은 금강 입구의 정점 G1에서 대부분의 계절동안 가장 낮은 밀도를 보였으며 외해역인 정점 G4로 나오면서 밀도가 증가하는 양상을 보였다(Fig. 5). 2017년 봄철과 가을철에는 정점 G1에서 2,080 ind./m2의 높은 밀도를 보였는데 옆새우류인 Sinocorophium sinensis의 밀도가 1,220 ind./m2로서 59%의 높은 점유율을 보였다. 정점 G5에서도 2,270 ind./m2를 보였는데 다모류인 Scoletoma longifolia 873 ind./m2, 이매패류인 Theora fragilis 467 ind./m2로서 각각 38% 및 21%의 점유율을 보였다. 조사 기간동안 가장 높은 밀도를 보인 정점은 2016년 8월의 G3으로서 5,537 ind./m2가 출현하였는데 이 시기는 옆새우류인 Gammaropsis utinomi가 22%, 다모류인 Amphicteis sp.가 34%로서 높은 점유율을 보였다.
생체량은 계절에 따라 59~277 g/m2 범위였으며 2015년 겨울철 및 2016년 겨울철과 봄철에 270 g/m2 이상의 높은 값을 보였으나, 그 밖의 계절에서는 유사한 양상을 보였다(Fig. 3). 공간분포 양상을 보면, 2015년 겨울철에는 정점 G1과 G6에서 500 g/m2 이상이 출현하였는데, G1에서는 이매패류인 Potamocorbula amurensis가 90% 이상의 점유율을 보였으며, G6에서는 생체량이 큰 이매패류인 Anadara broughtonii가 출현하여 70%를 점유하였다. 2016년 겨울철과 봄철에도 정점 G1에서 1,000 g/m2 이상의 생체량을 보였는데, P. amurensis가 각각 96% 이상의 점유율을 보였다.
종 다양도는 2.4~3.8 범위였으며 2015년과 2016년에는 계절에 따른 차이가 거의 없었으나 2017년에는 겨울철에서 여름철까지 감소한 후 가을철에 증가하는 양상이었다(Fig. 3). 담수의 영향을 가장 많이 받는 정점 G1에서 가장 낮고 외해역으로 나가면서 점차 증가하는 양상을 보였으며 가장 외해역의 정점 G4 및 고군산군도 남측과 비안도 사이에 위치한 정점 G5 및 G6은 유사한 양상을 보였다(Fig. 5).
3.3.3 낙동강 하구역
낙동강 하구역의 계절변동은 2015년 겨울철에 가장 많은 출현종수를 보인 후 다소 감소하는 양상이었으며, 2016년에는 전 계절이 다른 해에 비해 상대적으로 적은 출현종수를 보였다(Fig. 3). 2017년에는 겨울철에서 봄철로 가면서 증가한 후 여름철과 가을철에 감소한 양상이었다. 마산만 가장 내측에 위치한 정점 N1에서 가장 적은 출현종수를 보였으며 거제도 북측에 위치한 정점과 낙동강 하구역에 위치한 정점으로 나오면서 증가하는 공간분포 양상을 보였다(Fig. 6). 정점 N1에서는 2016년 8월에 생물이 출현하지 않았으며 11월에도 2종만이 출현하였다. 대부분의 계절동안 10종 미만의 출현종수를 보여 가장 적은 출현종수를 보였다. 인접한 정점 N2에서도 다른 정점들에 비해 상대적으로 적은 출현종수를 보였다. 낙동강 하구역에 위치한 정점 N5, N6, N7에서는 상대적으로 많은 출현종수를 보였다.
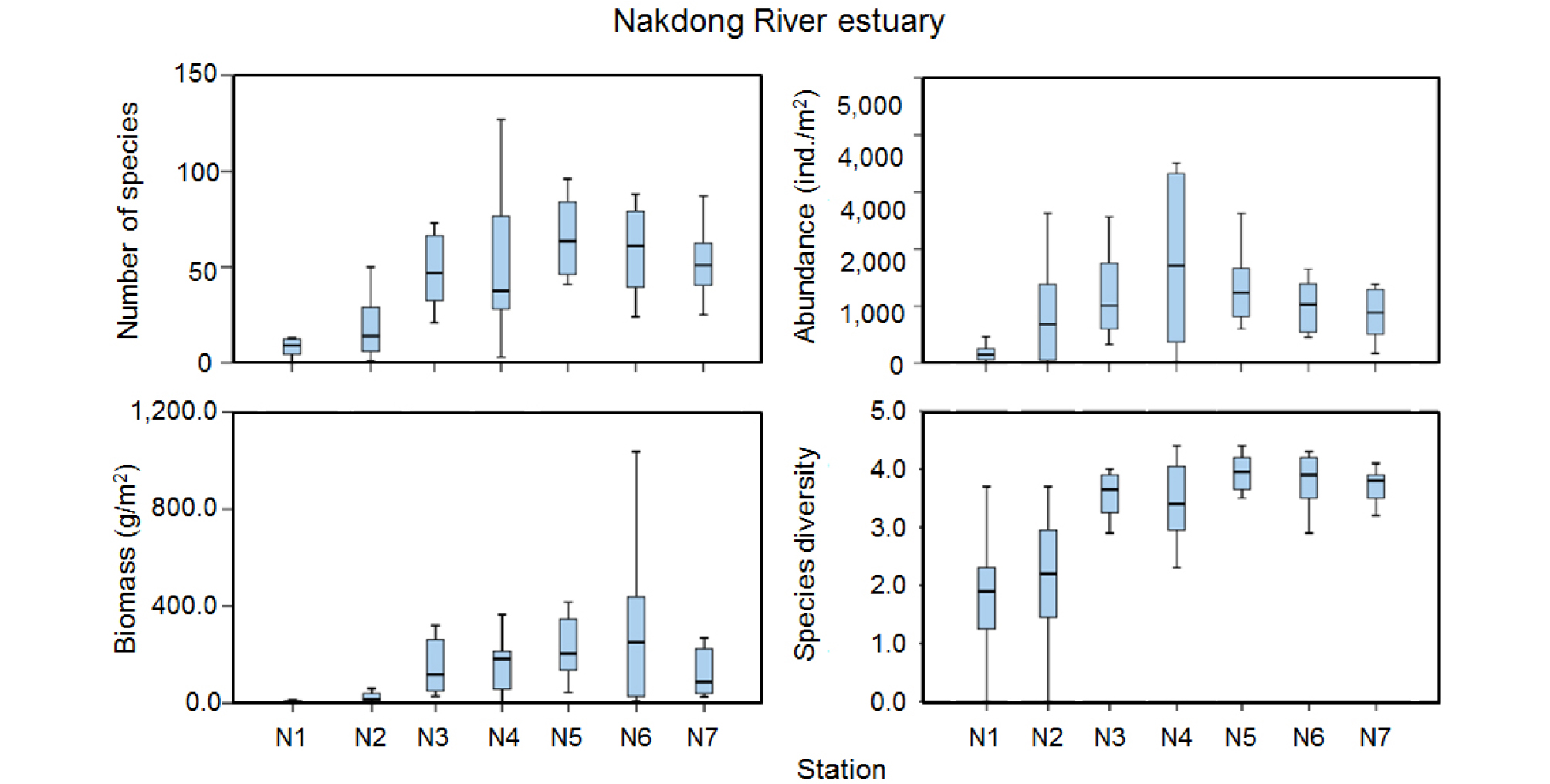
밀도는 2015년 겨울철에 2,000 ind./m2 이상이었으나 봄철로 가면서 감소한 후 여름철과 가을철에는 다소 증가하였다(Fig. 3). 2016년 겨울부터 2017년 봄철까지 1,000 ind./m2 미만의 밀도를 보이다가 2017년 여름철에 급격히 높은 밀도를 보였는데 정점 N4에서 40,000 ind./m2 이상이 출현하였으며 다모류 밀도가 96%를 점유하였다. 특히 다모류 가운데 Pseudopolydora sp.가 N4에서 출현한 밀도의 89%를 점유하였다. 공간분포 양상을 보면 만 내측의 정점 N1에서 가장 낮은 밀도였으며 외측으로 가면서 증가하는 양상을 보여 출현종수 공간분포 양상과 유사하였다. 또한 낙동강 하구역에 위치한 정점 N4에서는 계절적인 변동양상이 가장 크게 나타났다(Fig. 6).
생체량은 2015년과 2017년에는 겨울철에 상대적으로 낮고 가을철까지 지속적으로 증가하는 계절변동 양상을 보였다(Fig. 3). 그러나 2016년에는 겨울철부터 여름철까지 증가한 후 가을철에 감소한 양상이었다. 2015년 겨울철부터 2017년 봄철까지 극피동물의 생체량 점유율이 높아 60% 이상의 점유율을 보였으나 2017년 여름철과 가을철에는 연체동물의 생체량 점유율이 각각 74% 및 85%로 높았다. 공간분포 양상을 보면 출현종수 및 밀도 분포 양상과 동일하게 마산만 내측의 정점 N1 및 N2에서 낮고 외해역으로 나오면서 증가하는 양상을 보였다(Fig. 6). 이러한 출현종수 및 밀도의 시공간 변동 양상은 종 다양도에도 반영되어 나타났다.
3.4 주요 우점종
각 하구역에서 4% 이상의 밀도 점유율을 보이는 우점종은 Table 3에서와 같다. 한강 하구역의 경우 다모류인 Dispio oculata와 Heteromastus filiformis 및 Aonides oxycephala였으며, 금강 하구역에서는 다모류인 H. filiformis와 Scoletoma longifolia, 연체동물인 Potamocorbula amurensis 및 Theora fragilis였다. 또한 낙동강 하구역에서는 다모류인 Pseudopolydora sp., Aphelochaeta sp., Paraprionospio patiens, Magelona japonica와 이매패류인 T. fragilis였다.
Table 3. Abundance of dominant macrozoobenthic species in the three major estuaries of Korea, data pooled from February 2015 to November 2017
한강 하구역에서는 Dispio oculata의 경우 2016년 2월에 최대밀도를 보이고 5월에 다소 감소하였는데 그 밖의 계절에서는 출현하지 않거나 극히 낮은 밀도를 보였다(Fig. 7). 특히, 2016년 겨울철과 봄철에 정점 H4와 H6에서 각각 5,000 ind./m2 이상의 높은 밀도를 보였으며 그리나 다른 정점들에서는 출현하지 않거나 극히 낮은 밀도를 보였다. 따라서 이 종은 시공간적으로 제한적인 분포를 나타내는 특징을 보였다. 또한 H. filiformis는 전 계절에 걸쳐 대부분의 정점에서 출현하였다. 인천항 주변에 위치한 정점 H3에서 년 중 가장 높은 밀도로 출현하였으며, 가장 북측의 정점 H1과 시화호 방조제와 인접한 정점 H4에서는 밀도가 낮거나 계절적으로 출현하지 않는 특징을 보였다. 이 종은 기간이 경과함에 따라 전체적인 출현밀도가 다소 증가하는 양상을 보였다. A. oxycephala는 2015년 2월부터 11월까지 증가하다가 이후에는 출현하지 않거나 일부 정점에서 낮은 밀도로 출현하였다. 2015년 5월과 8월에는 외해역에 위치하여 모래 함량이 높은 정점 H6과 H7에서 2,000 ind./m2 이상의 밀도로 출현하였으며, 11월에는 정점 H6에서 8,793 ind./m2 의 높은 밀도를 보였다.
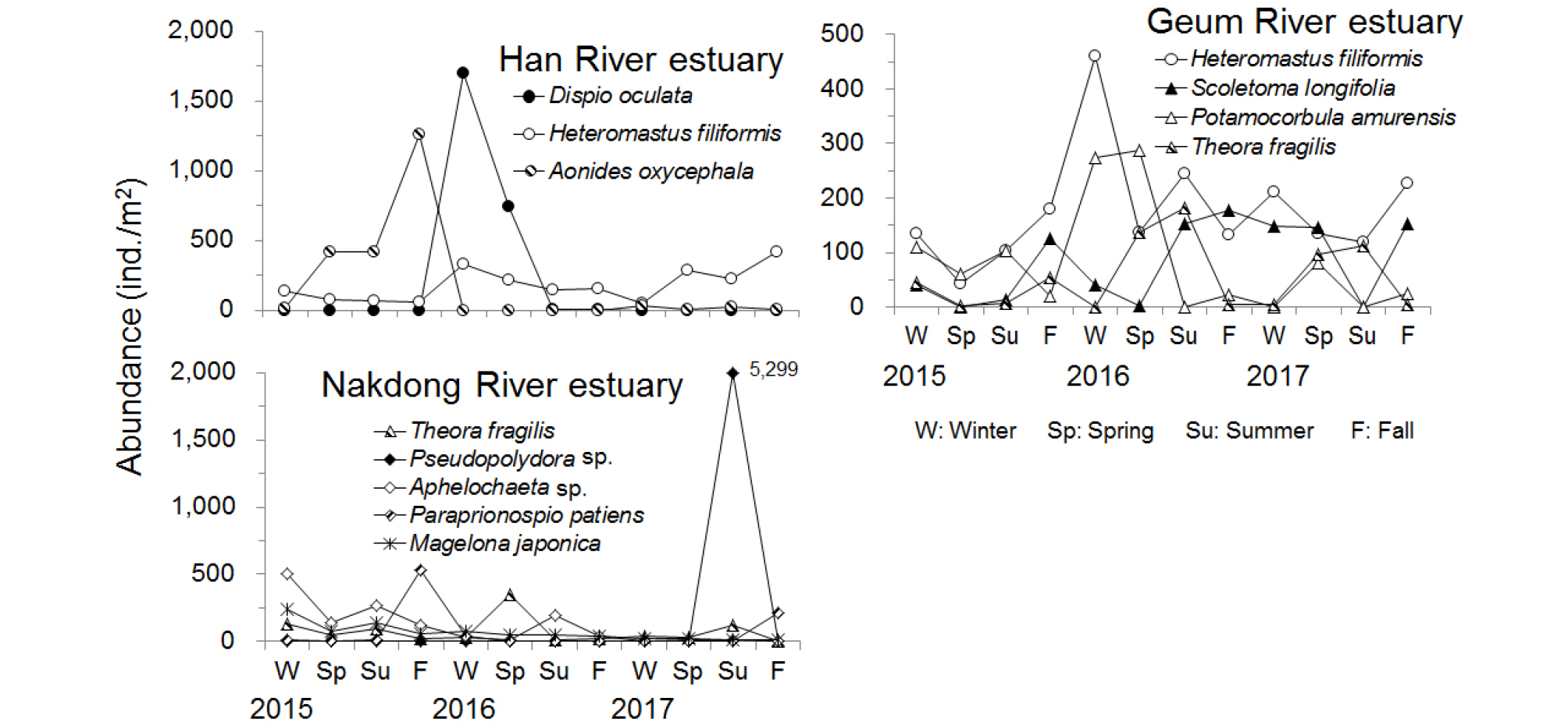
금강 하구역에서는 H. filiformis가 평균 밀도의 13%를 점유하여 가장 높은 비율을 차지하였다. 이 종은 2015년 봄철부터 증가하기 시작하여 2016년 2월에는 가장 높은 밀도를 보였으며 이후로는 계절에 따라 증가 및 감소 양상을 보였다(Fig. 7). 2016년 2월에는 정점 G3에서 1,710 ind./m2가 출현하여 이 정점에서 출현한 밀도의 76%를 점유하였으며, 전체적인 평균 밀도 증가에 기여하였다. S. longifolia는 6%의 점유율을 보였으며 2016년 8월부터 2017년 11월까지는 평균 100 ind./m2 이상의 밀도를 보였는데 2017년 8월에는 출현하지 않았다. 이 종은 대부분의 계절동안 정점 G5에서 높은 밀도로 출현하는 특징을 보였으며 정점 G5에서 출현 여부가 전체적인 밀도 증가에 영향을 미치고 있다. 금강 하구역 저서동물 군집의 가장 큰 특징으로서는 기수역에서 서식하는 P. amurensis가 세 번째 우점종으로 출현한 것을 들 수 있다. 이 종은 한강 하구역과 낙동강 하구역 조사정점에서는 출현하지 않은 종으로서 담수의 영향을 가장 많이 받는 정점 G1에서만 출현하는 특징을 보였다. 또한 유기물 오염지시종으로 알려진 T. fragilis는 4%의 점유율을 보였는데, 2016년 5월 및 8월에 높은 밀도를 보였다. 특히 두 계절 모두 정점 G5에서 각각 817 ind./m2 및 1,017 ind./m2가 출현하여 해당 정점에서 출현한 밀도의 각각 38% 및 37%를 점유하였다.
낙동강 하구역에서 가장 우점한 종은 다모류인 Pseudopolydora sp.였으며, 전체 평균 밀도의 약 28%의 점유율을 보였다(Table 3). 이 종은 2017년 8월에 가장 높은 밀도를 보였는데, 진해만 입구인 거제도 북측에 위치한 정점 N4에서 30,000 ind./m2 이상의 높은 밀도로 출현하여 이 정점에서 출현한 밀도의 90%를 점유하여 극우점하였다. 다모류인 Aphelochaeta sp.는 전체 평균 밀도의 7%를 점유하였으며, 2015년 2월과 11월에 상대적으로 가장 높은 밀도를 보였다(Fig. 7). 2월에는 정점 N4에서 3,467 ind./m2가 출현하여 이 정점에서 출현한 밀도의 42%를 점유하였다. 또한 T. fragilis는 마산만 내의 정점 N2 및 진해만 입구역의 정점 N3에서 주로 출현하는 특징을 보였다. 2016년 5월의 경우 정점 N2에서 2,127 ind./m2가 출현하여 해당 정점에서 출현한 밀도의 81%를 점유하였다. 그 밖에 다모류인 P. patiens 및 M. japonica도 주요 우점종으로 출현하는 특징을 보였다.
3.5 집괴분석
3.5.1 한강 하구역
한강 하구역의 경우 4개의 정점군으로 구분되었으며 이러한 정점군의 공간배치 양상은 계절에 따라 큰 차이가 없었다(Fig. 8). 영종도와 인천 사이에 위치한 정점 H1과 H2는 겨울철을 제외하고는 봄, 여름, 가을 모두 동일한 정점군으로 구분되었다(정점군 A). 또한 영종도 남측의 정점 H3은 4계절 모두 동일한 정점군(정점군 B)으로 구분되었다. 가장 외측의 정점 H6과 H7은 가을철의 정점 H7을 제외하고는 정점군 D로 구분되었으며, 시화방조제 앞 정점 H4와 무의도 남측의 정점 H5도 계절적으로 동일한 정점군에 포함되었다(정점군 C). 따라서 한강 하구역의 저서생태계는 갈수기인 겨울철에 담수의 영향을 가장 많이 받는 정점 H1을 제외하고는 전 계절에 걸쳐 정점군의 배치가 동일하게 유지되는 특징을 보였다. SIMPER분석 결과 한강 하구역의 각 정점군에서 출현한 우점종들은 영종도와 인천시 사이의 수로역에 위치한 정점군 A와 B의 경우 Heteromastus filiformis의 기여도가 크게 나타났으며, 시화 방조제와 무의도 남측의 정점군 C에서는 Aricidea jeffreysii, 가장 외해역에 위치한 정점군 D에서는 Aonides oxycephala의 밀도가 높게 나타났다(Table 4).
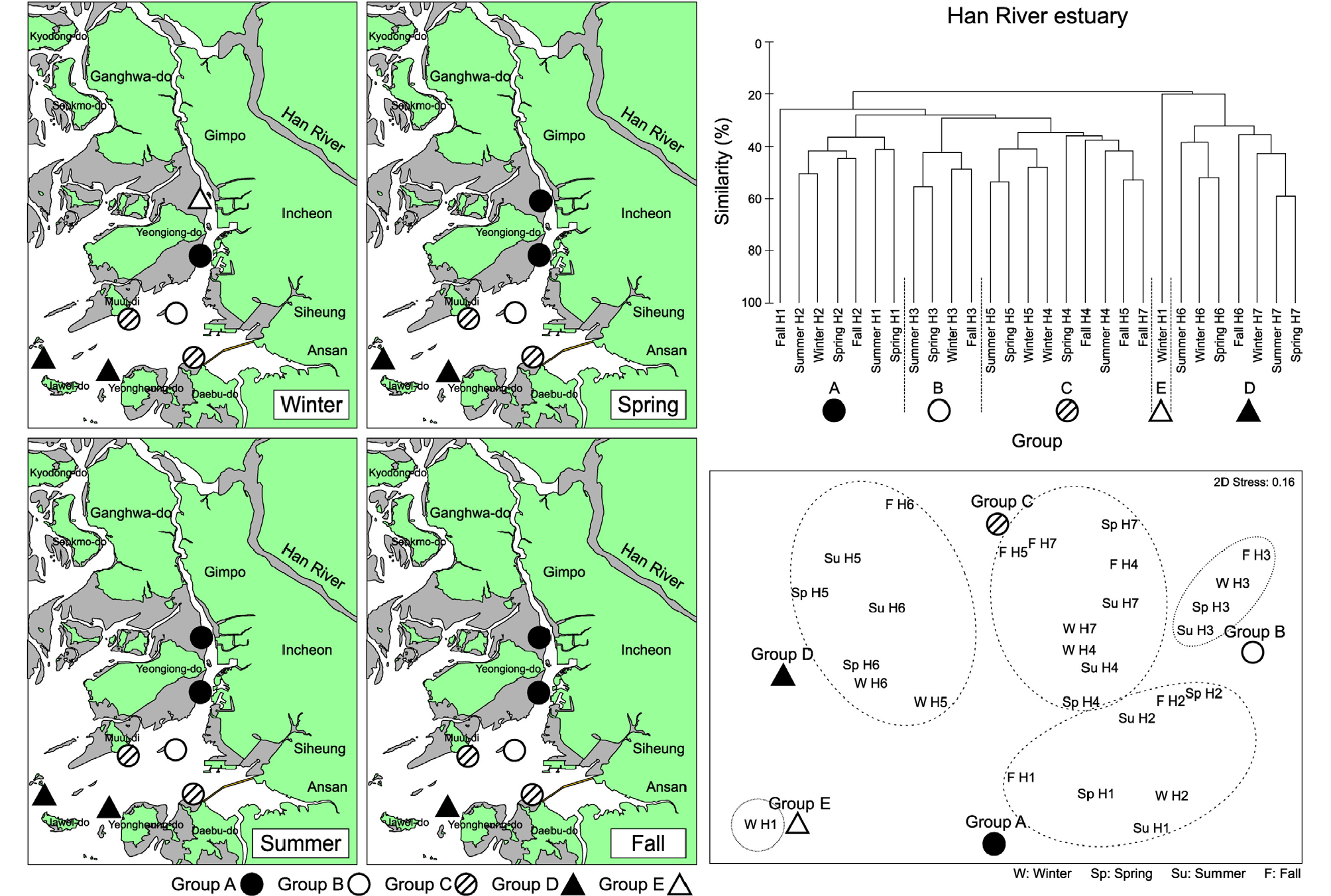
Table 4. Environmental and ecological characteristics in various station groups from Han, Geum and Nakdong Rivers
3.5.2 금강 하구역
금강 하구역의 경우 4개의 정점군으로 구분되었으며 이러한 정점군 공간 배치는 전 조사기간 동안 동일하게 나타났다(Fig. 9). 금강 하구역에 인접한 정점 G1부터 G4까지는 4계절 동안 별개의 정점군으로 구분되어 담수가 직접 유입되는 내측에서 외측으로 가면서 순차적으로 배치되는 특징을 보였다(정점군 A, B, C, D). 또한 고군산군도 남측의 새만금 방조제 인근의 정점 G5와 G6은 별개의 정점군으로 구분되었다(정점군 E). 따라서 금강 하구역의 저서생태계는 4계절에 걸쳐 담수의 영향을 가장 많이 받는 정점 G1부터 외해역의 정점 G4까지 순차적인 정점군 배치가 동일하게 유지되며, 고군산군도 남측의 정점군은 북측의 정점군과 분리되는 특징을 보였다. SIMPER분석 결과 금강의 경우 담수의 영향을 가장 많이 받는 정점군 A의 경우 Potamocorbula amurensis가 22%의 기여율을 보였다. 정점군 B와 C에서는 H. filiformis 가 각각 25% 및 10%의 기여율을 보였다. 정점군 D에서는 Nephtys polybranchia의 기여도가 약 6%로 높았으나 주요 우점종은 H. filiformis 및 Aphelochaeta sp.였다. 정점군 E에서는 Sternaspis scutata가 7%의 기여율을 보였으며, 주요 우점종은 Scoletoma longifolia 및 Theora fragilis였다(Table 4)
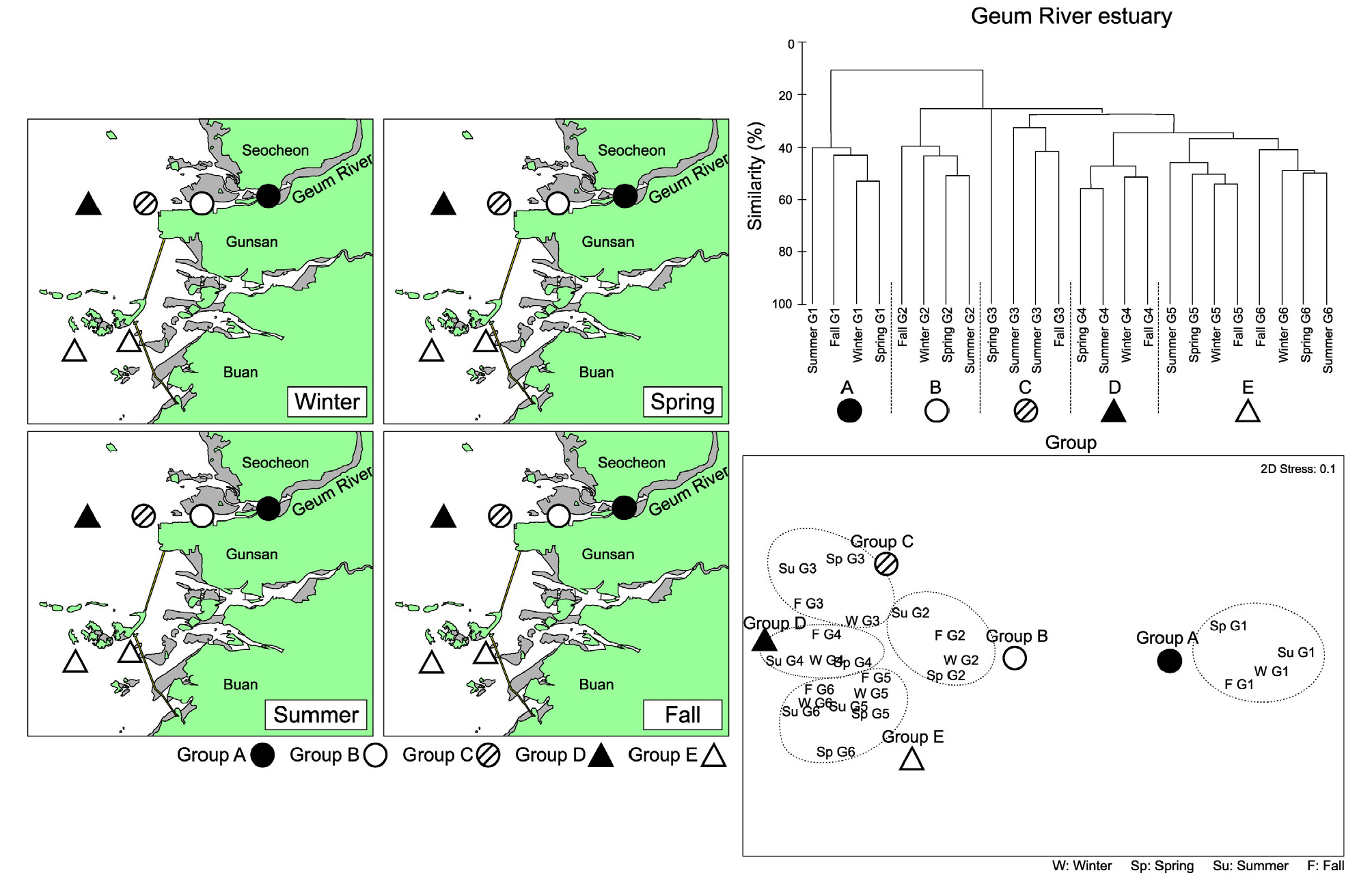
3.5.3 낙동강 하구역
낙동강 하구역은 4개의 정점군으로 구분되었는데 마산만 내측에 위치한 정점 N1과 N2는 겨울철과 봄철에는 동일한 정점군이었으나(정점군 A), 여름철에는 별개의 정점군으로 구분되었다(정점군 B). 가을철에도 가장 내측의 정점 N1과 인접한 정점 N2는 각각 서로 다른 정점군으로 구분되는 특징을 보였다(Fig. 10). 진해만 입구역에 해당하는 정점 N3과 N4는 정점군 C로 구분되었으며, 낙동강 하구역의 정점들(정점 N5, N6, N7)은 정점군 D로 구분되었다. 따라서 낙동강 하구역의 저서생태계는 담수 영향을 다소 받으면서 외해역으로 노출되어 있는 정점군 D와 진해만 입구 정점군 C, 계절적인 변동이 나타나는 마산만 내측의 정점군 A와 B로 구분되는 특징을 보였다. SIMPER분석 결과 마산만 내측의 정점군 A에서는 T. fragilis의 기여도가 27%로 높았으며, Paraprionospio patiens가 우점종으로 출현하였다. 정점군 B에서는 H. filiformis의 밀도가 높아 14%의 기여율을 보였으나 특징적인 우점종은 없었다. 정점군 C에서는 T. fragilis가 약 6%의 상대적으로 낮은 기여도를 보였다. 정점군 C에서는 Pseudopolydora sp.가 우점종으로 출현하였으나 출현 정점수가 극히 제한되어 있어 전제적인 정점군 구분에는 기여도가 낮았다. 정점군 D의 경우 Magelona japonica가 우점종으로 출현하였으며 약 6%의 기여도를 보였다(Table 4).
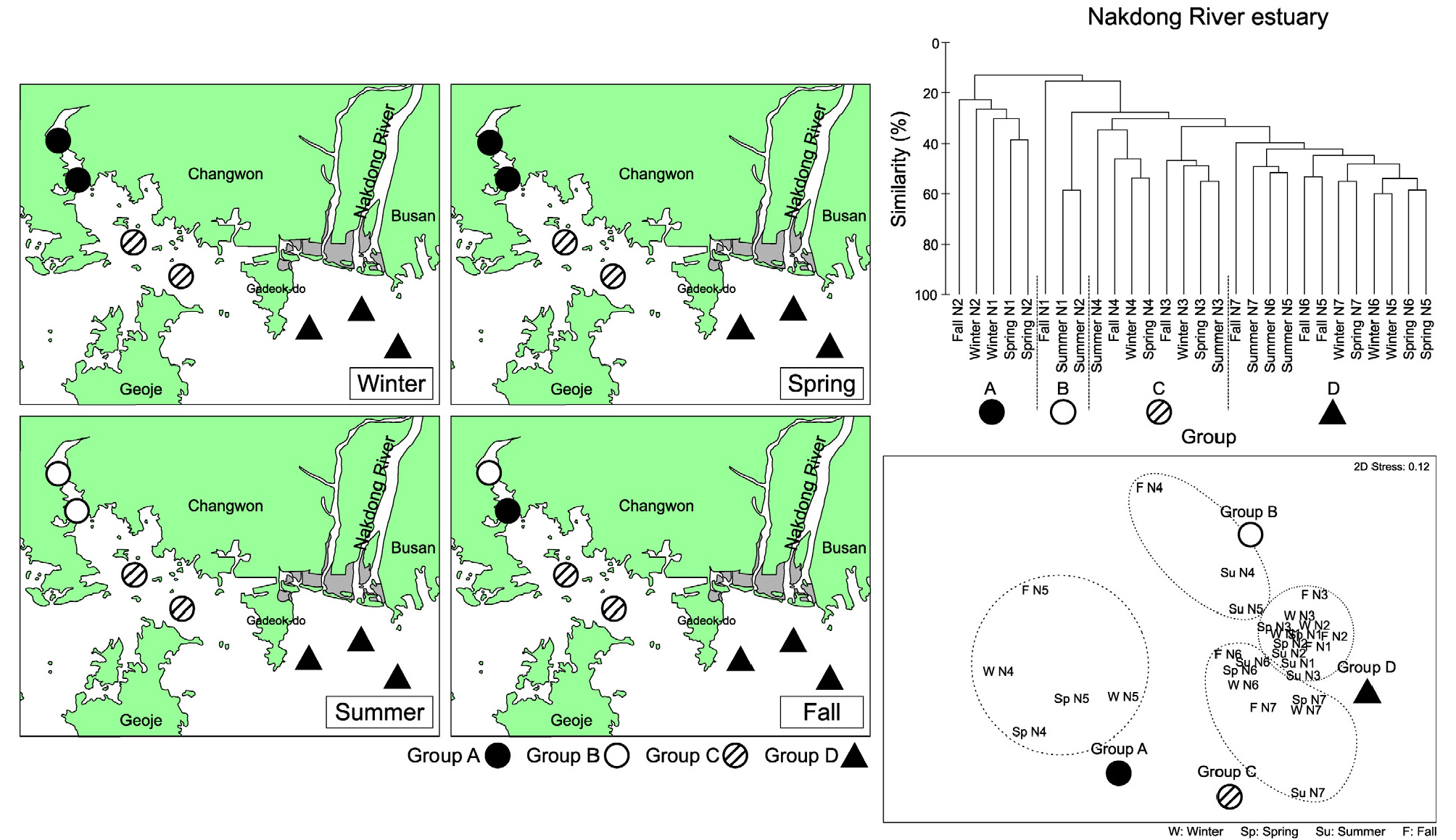
3.6 환경요소와의 관계
저서군집 요소에 영향을 미치는 환경요소는 각 하구역에 따라 다소 차이가 있었다(Tables 5, 6). 한강 하구역의 경우 조사 정점이 담수가 유입되는 입구로부터 떨어져 있어 염분에 의한 영향보다는 평균입도에 의해 영향을 받고 있었으나 낮은 상관관계를 보였다. 금강 하구역의 경우 염분과 실트함량 차이에 의해 군집이 영향을 받고 있었으며, 낙동강 하구역의 경우 마산만 내측에 위치한 정점으로 인해 용존산소와 염분, 퇴적상과 유기물 함량에 의해 복합적으로 군집 요소가 영향을 받고 있었다.
Table 5. Relationship between environmental variables and macrozoobenthic assemblage data using BIOENV analysis with environmental variables best explaining the faunal pattern. Resulting values are weighted Spearman rank correlation coefficient (r). Bold values represent the highest correlation value for the best explanatory variables
Table 6. Spearman's correlation coefficient between benthic habitat conditions and community parameters and the abundance of dominant species
저서동물 군집 요소와 환경요소와의 상관관계에서도 한강 하구역의 경우 수질환경 요소와 퇴적환경 요소와의 상관관계가 유의하게 나타났으며, 금강 및 낙동강 하구역의 경우 염분과 퇴적환경 요소에 의한 영향이 유의하게 나타났다(Table 6).
4. 고 찰
4.1 공간분포 특징
한강 하구역에서 조사시기동안 출현종수는 겨울철에는 상대적으로 낮고 봄철에 증가하여 여름철에 최대값을 보인 후 가을철에 다시 감소하는 매년 반복적인 양상을 보였다. 또한 영종도와 인천 사이의 수로역과 무의도 남측 정점에서 높은 출현종수를 보였다. 이러한 계절 변동과 공간분포 양상은 Lim et al.(1995)이 영종도 일대 18개 정점에서 4계절 동안 조사하고 이 일대의 대형저서동물은 여름철에 가장 많은 출현종수를 보였으며 영종도와 인천 사이의 수로역과 무의도 남측 정점에서 높은 출현종수를 보고한 결과와 유사하다. 또한 Shin et al.(1989, 1992)도 경기만 일대의 대형저서동물 공간분포 조사에서 이질적인 퇴적상을 가진 수로역에서 높은 출현종수를 보고하였다. 본 조사에서도 조사 정점수는 적지만 기존 연구 결과와 유사한 계절변동 및 공간분포 양상을 보이고 있다.
금강 하구역의 경우 담수가 유입되는 정점 G1부터 외해역으로 나가면서 군집의 출현종수 및 밀도가 증가하는 뚜렷한 경사를 보였다. 이는 염분 경사가 매우 뚜렷한 우리나라 소형 하천 자연 하구역에서의 대형저서동물 군집 공간분포 양상과 유사하였다. 금강은 해양에 인접하여 하구둑이 건설되었지만 바다로부터 약 10 km 상류에 위치하고 있어 하구둑 외측은 어느 정도 자연 하구의 특징이 남아 있는 장소로 볼 수 있다. 특히 우리나라 소형 하천 자연 하구역에서 특징적으로 서식하는 Potamocorbula amurensis가 정점 G1에서만 출현하는 것은 담수 유입으로 인해 기수역이 존재한다는 것을 의미한다. 기수역의 존재여부는 염분 농도에서도 파악되고 있다. 금강 하구역의 경우 가장 상부에 위치한 정점 G1의 경우 Venice system(1959)의 구분인 다함수역(polyhaline)에 속하는 22.1~28.8 psu의 저층염분 농도를 보였는데 30.0 psu 이상의 농도를 보인 다른 정점들에 비해 상대적으로 낮은 값이었다.
낙동강 하구역의 큰 특징은 해양에 인접하여 하구둑이 건설되어 있으므로 기수역의 특성이 사라졌다는 점이다. 따라서 하구둑 바깥쪽으로는 해양 환경이며 기수종이 출현하지 않고 있다. 하구둑 내측은 재첩류(Corbicula sp.)와 같은 전형적인 기수종들이 출현하는 것으로 보고되어 있으나 본 조사는 외측 해역에서만 이루어졌기 때문에 확인이 불가능하였다. 그러나 낙동강 하구역은 하구둑이 건설된 이후 내측의 담수종과 외측의 해수종으로 구분되는 생태계 특징을 보이는 것으로 알려져 있어(Park et al., 2016), 담수의 유입이 자유로울 경우 전형적인 하구역 생태계가 형성될 수 있다. 본 조사 해역인 마산만 내측의 정점에서는 대형저서동물 군집이 가장 빈약하였으며, 외해역으로 가면서 출현종수와 종 다양도가 증가하는 양상이었다. 특히 마산만 내측의 경우 여름철 빈산소수괴의 형성이 대형저서동물 군집을 빈약하게 하는 원인으로 판단되며, 이러한 현상은 Lim et al.(2006)에 의해 상세히 보고되었다. 해양수산부 수질자동측정망 자료에 따르면 마산만 내측은 4월부터 빈산소환경이 형성되어 9월~10월까지 지속되며, 이는 Lim et al.(2006)의 결과에서도 확인된다. 빈산소수괴가 소멸된 후 저서동물 군집은 2~3개월 이후에 마산만 외측에서 내측으로 가면서 점차 회복되는 것으로 보고되어 빈산소수괴 형성이 대형저서동물 군집에 심각한 영향을 미치고 있음을 알 수 있다. 마산만 내측에서 회복된 저서동물 군집은 이듬해 빈산소수괴 형성으로 집단 폐사하여 여름철 마산만 내측 수질악화의 원인을 제공하고 있는 것으로 보인다(Cha, 2015).
각 하구역의 대형저서동물 군집 요소의 공간분포 양상은 하구역에 따라 다소 차이가 있었으나 각 하구역의 환경 특성을 잘 나타내주고 있다. 한강 하구역에서는 염분에 의한 영향보다는 평균입도와 분급에 의해 영향을 받는 것으로 나타났는데 이것은 각 조사 정점이 담수가 유입되는 입구로부터 떨어져 있어 담수의 영향이 약하기 때문으로 보여진다. 실제로 한강 하구역 각 정점들에서의 저층 염분 분포는 계절적으로는 다소 차이가 있으나 공간적으로 큰 차이가 없었다. 또한 외해역으로 나가면서 퇴적물의 모래 함량이 높고 입도가 조립해지면서 분급이 큰 이질적인 퇴적환경을 나타내고 있다. 이러한 저서 환경 특성은 Shin et al.(1989)과 Lim et al.(1995)에 의해 보고된 바도 있다. 따라서 한강 하구역의 경우 저서생태계를 보호하기 위해서는 이 해역에서의 퇴적환경 변화를 초래하는 요인들(골재채취, 제방축조, 매립 등)에 대한 관리가 우선되어야 할 것으로 보인다. 또한 금강 하구역의 경우 염분과 모래 함량 차이에 의해 군집이 영향을 받고 있어 금강으로부터의 담수유입 및 주변 퇴적 환경에 변화를 줄 수 있는 요인들에 대한 관리가 우선시 되어야 할 것으로 보인다. 낙동강 하구역의 경우 염분과 저층 빈산소수괴 형성 및 세립한 퇴적상으로 인한 높은 유기물 함량에 의해 복합적으로 군집 요소가 영향을 받고 있어 마산만 내측의 빈산소수괴 및 유기물 함량에 대한 관리를 중점적으로 해야 할 것으로 판단된다.
4.2 주요 우점종의 조성 및 분포
조사해역에서 출현한 우점종은 하구역에 따라 차이가 나타났다. 3개 하구역에서 공통적으로 4% 이상의 점유율을 보이는 종은 없었으며, 우리나라 연안역에서 광범위하게 출현하는 것으로 보고된 Heteromastus filiformis는 한강 및 금강 하구역에서는 각각 16% 및 13%로서 높은 점유율을 보였으나 낙동강 하구역에서는 약 3.6%의 점유율을 보였다. 또한 Theora fragilis는 금강과 낙동강 하구역에서 공통적으로 각각 4% 이상의 분포를 보였다. 이들 두 종은 입도가 세립하고 유기물 함량이 높은 우리나라 연안역에서 주요 우점종으로 광범위하게 출현하는 종으로 알려져 있다. 특히 T. fragilis는 우리나라의 경우 영산강 하구역(Lim and Park, 1999)이나 진해만(Lim and Hong, 1997) 뿐만 아니라 일본에서도 오래전부터 부영양화된 해역의 지표종으로 사용되어 왔다(Imabayashi and Tsukuda, 1984). 따라서 금강과 낙동강 하구역에서 이 종이 높은 밀도로 출현하는 것은 펄 함량이 높고 입도가 상대적으로 세립한 이 두 해역의 저서환경과 밀접한 연관성을 가지고 있는 것으로 판단된다.
한강 하구역에서 가장 우점적으로 출현한 Dispio oculata는 우리나라 다른 해역에서는 보고된 바가 없는 종이다. 이 종은 영흥도 북측의 정점 H4와 H6에서만 일시적으로 5,000 ind./m2 이상의 높은 밀도를 보였으며 다른 정점들에서는 출현하지 않거나 극히 낮은 밀도를 보임으로서 제한적인 분포 특성을 보였다. D. oculata는 Imajima(1990)에 의해 일본 Wakasa Bay의 Yura 강 하구역에서 신종으로 처음 보고된 종으로서 수심 5m 이하의 연안에서 주로 출현하며, Dispio 속(genus)에 속하는 종들은 대부분 강 하구역에서 서식하는 것으로 보고되고 있어 이 종의 서식처를 유추해 볼 수 있다. Aonides oxycephala는 표층퇴적물식자(surface deposit feeder)로서 밀도는 퇴적물의 펄 함량과 음의 상관관계를 가진 것으로 보고되고 있다(Thrush et al., 2003). 이 종은 2015년 5월과 8월에 외해역인 정점 H6과 H7에서 2,000 ind./m2 이상의 밀도로 출현하였으며, 11월에는 정점 H6에서 8,793 ind./m2의 높은 밀도를 보였는데 이들 정점은 모래 함량이 90% 이상으로 높게 나타났다. 이 종은 환경이 파괴되어 생물이 사라진 장소에 처음으로 성체가 가입하여 다른 종의 가입을 촉진하는 것으로도 알려져 있다(Thrush et al., 1992).
우리나라 연안에서 광범위하게 서식하는 H. filiformis가 한강 하구역에서도 광범위하게 출현하였다. 인천항 주변에 위치한 정점 H3에서 4계절 내내 가장 높은 밀도로 출현하는 특징을 보였으며, 가장 북측의 정점 H1과 시화호 방조제와 인접한 정점 H4에서는 밀도가 낮거나 계절적으로 출현하지 않는 특징을 보였다. 따라서 이 종은 담수의 영향이 강하거나 모래 함량이 많은 외해역의 정점들에서는 출현하지 않는 특징을 보였다. 특히 모래 함량이 높은 곳에 서식하는 A. oxycephala는 펄 퇴적과 같은 퇴적환경 변화를 모니터링 할 수 있는 지시종으로 이용할 가치가 있을 것으로 판단된다. 또한 D. oculata의 분포 양상으로 보아 영흥도 북측의 시화방조제 수문에 인접한 정점 H4와 H6에서는 여전히 담수의 영향을 받고 있으며 고유한 생태계가 형성되어 있음을 알 수 있다. 그러나 이들은 2016년 2월과 5월에 주로 출현하며 그 밖의 계절에서는 출현하지 않거나 극히 낮은 밀도이기 때문에 담수 유입과의 연관성이 있는지는 보다 세밀한 검토가 필요하다. 따라서 우점종의 출현양상으로부터 한강 하구역은 담수의 영향을 받고 있으며 퇴적물의 유기물 함량이 높다는 것을 잘 반영하고 있다.
금강 하구역의 경우, 퇴적물에 펄 함량이 높아 유기물 함량이 높은 장소에서 높은 밀도로 출현하는 H. filiformis가 전 계절에 걸쳐 대부분의 정점에서 높은 밀도로 출현하는 특징을 보였다. 한강 하구역의 모래 함량이 높은 외해역에서 높은 밀도로 출현하던 A. oxycephala는 출현하지 않아 입도가 세립한 것을 알 수 있다. 이것은 새만금 방조제 이후 조립질 퇴적상이 세립질 퇴적상으로 변하였다는 보고(An et al., 2006)와 일치하는 현상이다. 또한 Scoletoma longifolia도 두 번째 높은 밀도로 출현하여 이러한 퇴적환경 변화를 뒷받침하고 있다. S. longifolia는 2016년 8월부터 2017년 11월까지는 평균 100 ind./m2 이상의 밀도를 보였으나 2017년 8월에는 출현하지 않았는데 이후 다시 출현하는 것으로 보아 일시적으로 출현하지 않는 것으로 볼 수 있다. 이 종은 대부분의 계절동안 정점 G5에서 높은 밀도로 출현하는 특징을 보이는 종으로서 정점 G5에서 소멸될 경우 전체적으로 출현하지 않게 된다. 정점 G5에서의 어떠한 환경요소의 변화가 이 종의 일시적인 소멸을 초래하였는지에 대해 면밀한 검토가 필요하다. 금강 하구역에서 출현한 가장 큰 특징으로서는 기수역에서 서식하는 P. amurensis가 세 번째 우점종으로 출현한 것을 들 수 있다. 이 종은 담수의 영향을 가장 많이 받는 정점 G1에서만 출현하는 특징을 보였으며, 한강 하구역과 낙동강 하구역 조사정점에서는 출현하지 않은 종이다. 소형하천인 수어천 및 와탄천에서 높은 밀도로 보고되었으며, 금강 하구역의 경우 새만금 방조제가 건설되기 전에는 기수역의 특징적인 우점종으로 보고되었다(Choi and Koh, 1994). 금강 하구역의 정점 G1에서 이 종이 출현하는 것은 담수의 영향을 받는 기수역 생태계가 유지되고 있음을 나타낸다.
낙동강 하구역에서 우점종으로 출현한 Pseudopolydora sp.는 2017년 8월에 진해만의 입구인 정점 N4에서 30,000 ind./m2 이상의 높은 밀도로 출현하여 이 정점에서 출현한 밀도의 90%를 점유하여 극우점하였다. 특히 이 종은 H2S 함량이 높은 퇴적물의 표층에 높은 밀도로 서식하는 것으로 보고되어 있다(Kanaya et al., 2016). 정점 N4에서 일시적으로 높은 밀도로 출현한 이 종은 기존 조사(Lim and Hong, 1997; Lee et al., 2005)에서는 보고되지 않은 종이었다. Seo et al.(2016)에 의하면 낙동강 하구둑에서 방류를 하면 낙동강 입구 정점에서 일시적으로 Pseudopolydora kempi의 유생이 대량으로 유입되어 정착하여 서식하다가 저층환경(퇴적상, 유기물 함량)으로 인하여 개체군 유지를 못하고 폐사한다고 보고되었다. 본 논문에서는 속명으로만 기재되어 있어 종 동정에 대한 면밀한 추가 조사가 있어야 하겠지만 낙동강 하구에서 일시적으로 우점하는 경향으로 보아 동일한 종으로 판단되면 향후 면밀한 감시가 필요하다. Lee et al.(2005)은 낙동강 하구역 일대에 대한 여름철 1회의 대형저서동물 군집조사에서 우점종으로 Tharyx sp., S. longifolia, T. fragilis를 보고하였다. 이것은 본 조사에서 출현한 Aphelochaeta sp. 및 T. fragilis 와는 일치하지만 다른 우점종과는 차이가 있었다. 이것은 이번 조사 해역은 마산만 내측까지 조사된 결과로서 조사장소의 범위 차이 때문으로 보여진다. 한편, Lim and Hong(1997)은 진해만 중앙부에서는 Paraprionospio pinnata가 우점종으로 출현하였으며 진해만 입구역(현재의 정점 N4 주변)에서는 Magelona japonica가 우점종으로 출현함을 보고하였다. Yokoyama and Choi(2010)는 우리나라에서 보고된 P. pinnata를 이후 P. coora, P. patiens 및 P. cordifolia로 구분하였기 때문에 정점 N2에서 출현한 우점종인 P. patiens는 과거 P. pinnata로 동정되었던 종의 출현 양상과 유사하다. 또한 가덕도 동측의 낙동강 담수의 영향을 가장 크게 받는 정점들에서 높은 밀도로 출현하는 M. japonica도 Lim and Hong(1997)의 결과와 유사한 분포양상을 보였다. 이러한 결과는 낙동강 하구역은 시간 경과에 따른 우점종 종류의 변화보다는 환경 변화에 따른 우점종의 분포역이 달라진다는 것을 의미한다. 따라서 마산만의 빈산소수괴 발달 양상과 우점종의 공간 분포 범위에 대한 면밀한 모니터링이 필요하다.
4.3 군집 구조
한강, 금강, 낙동강 하구역의 대형저서동물 군집의 공간분포 특징은 한강 하구역과 마산만 내에 위치한 일부 정점들을 제외하고는 전 계절동안 변화가 없이 동일한 정점군 분포를 보인다는 점이다. 한강 하구의 경우 한강으로부터 유입되는 담수의 영향을 가장 크게 받는 정점 H1은 다른 계절과 달리 겨울철에는 별도의 정점군으로 구분되는데 빈약한 저서군집을 보였다. 정점군 D의 경우에도 가장 외해역의 정점 H7이 가을철에 내측의 정점군 C에 속하는 것을 제외하고는 계절적으로 동일한 정점군으로 구분되어 안정된 양상을 보였다. 특히 정점 H3은 전 계절동안 동일한 정점군으로 구분되었다. 따라서 가장 북측의 정점 H1과 외해역의 정점 H7의 경우에만 정점군 구분이 계절적으로 다소 차이가 나타나지만 전체적으로는 변동이 거의 없었다.
금강 하구역의 경우에도 고군산군도 북측의 금강 하구역에 위치한 4개 정점들은 담수의 영향을 직접 받는 정점 G1부터 외해역에 이르는 정점 G4까지 전 계절을 통해 동일한 정점군 배치를 보인다. 또한 고군산군도 남측의 정점들도 전 계절을 통하여 별도의 정점군으로 구분되었다. 따라서 금강 하구역은 금강으로부터 유입되는 담수에 의한 염분의 차이가 저서동물 군집의 공간분포에 큰 영향을 미치고 있는 것으로 추정된다.
낙동강 하구역의 경우, 특히 마산만 내에 위치한 정점 N1과 N2에서는 계절에 따른 환경 변화를 뚜렷하게 반영하고 있다. 즉, 마산만 내측은 우리나라에서 저층 빈산소 수괴가 형성되는 대표적인 장소로 알려져 있는데(Lim et al., 2006), 표저층수괴의 혼합이 제한되는 여름철의 경우 다른 계절과는 달리 별개의 정점군으로 뚜렷하게 구분된다. 표저층수의 혼합이 일어나기 시작하는 가을철이 되더라도 내측에 위치한 정점 H1은 여름철의 군집 배치가 그대로 유지되며 겨울이 되면 정점 N1과 N2가 동일한 정점군으로 구분된다. 이 장소에서의 용존산소 변화를 보면 여름철에 가장 낮아지며 가을철에는 회복되는 계절변화를 보인다. 이와 같은 저층용존산소 농도 변화에 따라 정점군이 변하는 것은 Lim et al.(2006)에서 상세히 보고되어 있으며 본 조사결과도 일치하는 양상을 보였다.
5. 결 론
하구역에는 다양한 산업시설이 발달하고 도시가 발달하며 이로 인해 해양환경이 훼손되는 경우가 전세계적으로 나타나고 있다. 우리나라의 한강, 금강, 영산강, 섬진강, 낙동강 하구역에는 예외없이 산업시설과 도시가 발달하여 오염의 영향을 받고 있다. 더구나 금강, 영산강, 낙동강 하구역은 교통 편의 및 토지개발을 위해 하구둑을 막아 하수와 담수의 자연적인 혼합을 차단함으로서 하구역의 기능을 상실한 장소이다. 이러한 하구역의 생태계를 보호하고 관리하기 위해서는 지속적인 모니터링이 수행되어야 한다. 이러한 저서생태계 구조는 현재의 하구역 환경이 변할 경우 변동할 수 있으므로 본 결과는 하구역 관리에 중요한 참조 자료(reference data)로 사용할 수 있으며, 기초적인 모니터링 자료 축적이라는 데 의미가 있다.
본 연구 결과 각 하구역 저서동물 군집의 시․공간 변동양상에 영향을 미치는 환경 요소는 각각 다르게 나타났다. 이러한 결과로부터 한강 하구역의 경우에는 퇴적환경 변화를 초래하는 요인들(골재채취, 제방축조, 매립 등)에 대한 관리가 우선되어야 할 것으로 판단되며, 금강 하구역에서는 금강으로부터의 담수유입 및 주변 퇴적 환경에 변화를 줄 수 있는 요인들에 대한 관리가 우선시 되어야 할 것으로 보인다. 또한 낙동강 하구역의 경우 염분과 저층 빈산소수괴 형성 및 세립한 퇴적상으로 인한 높은 유기물 함량에 의해 복합적으로 군집 요소가 영향을 받고 있었다. 따라서 마산만 내측의 빈산소수괴 발달양상과 주요 우점종의 공간분포에 대한 모니터링을 중점적으로 해야 할 것으로 판단된다.
