

1. 서 론
2. 재료 및 방법
2.1 거머리말 이식
2.2 생육 환경 조사
2.3 이식 거머리말 조사
2.4 통계 분석
3. 결 과
3.1 생육 환경
3.2 이식 거머리말
4. 토 론
1. 서 론
해양현화식물인 잘피는 전 세계 연안에 약 60 여종이 서식하고 있으며, 우리나라에는 9종이 서식하고 있다(Short et al., 2007; Kim et al., 2012). 잘피서식지는 다양한 해양동물들에게 먹이, 서식처와 산란장을 제공하여 연안의 생물다양성을 향상시킨다(Hovel et al., 2002). 또한, 잘피는 잎과 엽초로 해수 중의 영양염을 흡수하고 지하경과 뿌리로 저질을 안정화시키고 퇴적물의 재부유를 감소시켜, 연안의 수질 향상과 환경 정화의 기능을 발휘한다(Thomas and Cornelisen, 2003). 그러나 최근 수십 년간 매립, 해안 개발과 급격한 부영양화 등 인위적 요인으로 전 세계 뿐만 아니라 우리나라의 잘피서식지는 급격히 감소하여 약 50%이상이 훼손되었다고 보고되고 있다(Short and Wyllie-Echeverria, 1996; Park and Lee, 2007). 연안생태계에서 잘피서식지의 생태적 가치가 재조명되면서 훼손된 잘피서식지를 복원하고 보전하려는 시도들이 활발히 진행되고 있다(Fonseca et al., 1994; Paling et al., 2001).
훼손된 잘피서식지를 복원하는 방법으로는 환경을 개선하여 잘피군락이 회복되기를 기다리는 소극적인 방법과 잘피 이식 등의 적극적인 방법으로 구분할 수 있다. 환경을 개선하는 방법은 잘피가 사라진 해역을 잘피생육에 적합한 수질환경으로 조성하여 자연적인 잘피 회복을 기대할 수 있지만 너무나 긴 시간이 소요되는 경향이 있다(Seddon, 2004). 따라서, 건강한 잘피묘를 이식하거나 종자를 파종하여 잘피군락지를 조성하는 방법이 1980년대 이후부터 미국, 호주, 일본 등에서 본격적으로 이용되어 왔다(Fonseca et al., 1994; Davis and Short, 1997; Paling et al., 2001). 우리나라에서도 최근 수산자원 회복을 위한 바다숲 조성사업에서 거머리말 성체 이식으로 거머리말 서식지를 조성하는 방법이 빈번하게 활용되고 있다(FIRA, 2019).
잘피서식지를 성공적으로 복원하거나 조성하기 위해서 잘피에 적합한 이식 방법과 장소의 선정이 중요하다. 잘피의 적정한 이식 방법은 대상 잘피 종과 이식 장소의 환경에 따라 달라 질 수 있다. 거머리말은 우리나라를 포함한 북반구에서 가장 넓은 서식면적을 가지고 있으며 동시에 서식지 훼손이 가장 많이 발생한 거머리말 종으로 대표적인 잘피서식지 복원의 대상으로 알려져 왔다(Fonseca et al., 1994; Short et al., 2002; Park and Lee, 2007). 거머리말은 철사고정법, 이식망법, 패각투척법, 점토한지법, 대나무 꽂이법 등 다양한 이식방법들이 개발되었다(Park and Lee, 2007; FIRA, 2019). 그 중 철사고정법은 다양한 퇴적물 상태에서 높은 생존율을 보이며, 새로운 환경에 적응하는 착생기간이 짧아 생육 밀도의 증가가 빠르다고 보고된 이식 방법으로 우리나라에서도 널리 활용되고 있다(Park and Lee, 2007; Park et al., 2009, 2011, 2013).
적정한 이식 방법과 함께 적합한 잘피 이식 장소의 선정은 성공적인 잘피 이식에 결정적인 영향을 미친다(Short et al., 2002; Park and Lee, 2007). 적합한 이식 장소를 찾기 위해 잘피 생육에 충분한 광량, 조류 및 파도의 세기, 수심, 과거 잘피의 생육 여부와 생물교란 정도 등을 종합적으로 평가하여 이식 후보 장소로 선정한다(Short et al., 2002). 잘피 생육이 가능한 후보 장소라 하더라도 지협적인 환경이나 경합생물과의 경쟁으로 인해 잘피 군락지 형성 가능성이나 속도는 달라질 수 있다(Park et al., 2013). 잘피군락지는 태풍, 허리케인이나 사이클론과 같은 강력한 태풍의 경로에 놓이게 되어 피해를 입거나, 예상하지 못한 긴 홍수로 인한 과도한 담수의 유입으로 저염수에 노출되어 훼손되기도 한다(Kim et al., 2015). 그래서 안정적인 잘피군락지 조성을 위해 후보 장소에서의 예비 이식 실험이 더욱 요구된다. 예비 이식 실험은 이식 후보 장소에 예비적으로 잘피를 이식하여 잘피의 생존율이 50% 이상 확보되고, 이식된 잘피가 번식하여 새로운 개체들이 생성되고 안정적으로 생장하여 정착하는 것을 확인하는 과정이다(Park et al., 2009, 2011, 2013). 따라서 잘피의 예비 이식 실험은 본격적인 잘피서식지 복원과 대규모 잘피 군락 조성의 위험성을 줄이고 성공률을 높일 수 있는 과정으로 볼 수 있다.
우리나라의 가장 큰 섬인 제주도는 동북 연안을 중심으로 거머리말이 자생하고 있다(Park et al., 2012). 제주도 동쪽에 위치한 방두만은 과거 만의 동쪽 해안을 중심으로 거머리말 군락지가 형성되어 있었으나, 현재는 모두 소실되었다. 본 연구에서는 방두만의 잘피서식지 복원을 위한 예비 이식 실험으로 2019년 1월과 2020년 1월말에 내만 6개 이식 후보 장소에 철사고정법을 이용하여 잘피를 이식하였다. 그 후 약 10개월간 각 장소의 환경변화 즉, 수중광량, 수온, 염분 농도와 거머리말을 뒤덮어 광합성을 제한할 수 있는 부유성 해조류인 파래의 누적 높이와 함께 이식 잘피의 밀도, 생존율과 형태적 특성을 조사하였다. 본 연구 결과는 방두만의 거머리말 이식실험을 통해 서식지 복원 가능성을 검토함으로써, 효율적인 잘피 군락 복원을 위한 이식적지를 선정하는데 귀중한 자료를 제공할 것이다.
2. 재료 및 방법
2.1 거머리말 이식
거머리말 이식 실험 장소인 방두만은 제주도 동쪽 신양섭지해수욕장 주변 해안에 위치한다(Fig. 1). 방두만의 북쪽과 동쪽에는 넓은 모래사장이 있으며 만 내부의 평균 수심은 약 3 m이고 표층 퇴적물은 사질로 이루어져 있다(Hwang and Koh, 2012). 방두만은 형성 초기에는 개방형 만이었으나 1990년대 후반, 만의 서쪽 입구에 방파제가 건설되어 반폐쇄적 내만의 형태를 이루고 있으며 입구의 폭은 약 300 m이다. 만의 서쪽에는 육상양식장 시설이 위치하고 북서쪽 신양 포구에서는 지하수가 용출되며 2000년대 초부터 여름철 부유성 해조류인 갈파래의 과잉 증식 현상이 보고되고 있다(Song et al., 2016). 방파제 건설 전 만의 동쪽 해안을 중심으로 거머리말 군락지가 비교적 넓게 펼쳐져 있었으나 모두 소실되었고, 현재는 만 서쪽 입구 항내에만 소규모 거머리말 군락지가 남아 있다. 본 실험의 거머리말 이식은 2019년 1월말과 2020년 1월말에 걸쳐 방두만 거머리말 이식 후보지 6개소에서 실시하였다. 2019년 수심의 차이가 있는 St. 1과 St. 2을 이식 후보지로 비교하였고, 2020년 이식 거머리말의 높은 생존율 및 생장성을 보이는 St. 1과 유사한 수심대에 위치하면서 갈파래의 누적 높이가 다른 St. 3 ~ St. 6으로 후보지를 확장하였다(Table 1). 이식에 사용된 공급용 거머리말은 모두 제주시 조천읍 조천항에서 채취하였으며, 채취된 개체는 지하부의 퇴적물을 제거한 후 해수가 채워진 플라스틱 용기에 담아 이식 장소로 옮긴 후 2일 이내 각 장소에 식재하였다. 거머리말 이식 방법은 약 15 cm 길이의 v자형 철사에 4마디 이상의 지하경을 가진 거머리말 6개체씩을 40 cm 간격으로 식재하는 철사고정법을 이용하여 각 장소 당 약 500개체의 잘피를 이식하였다.
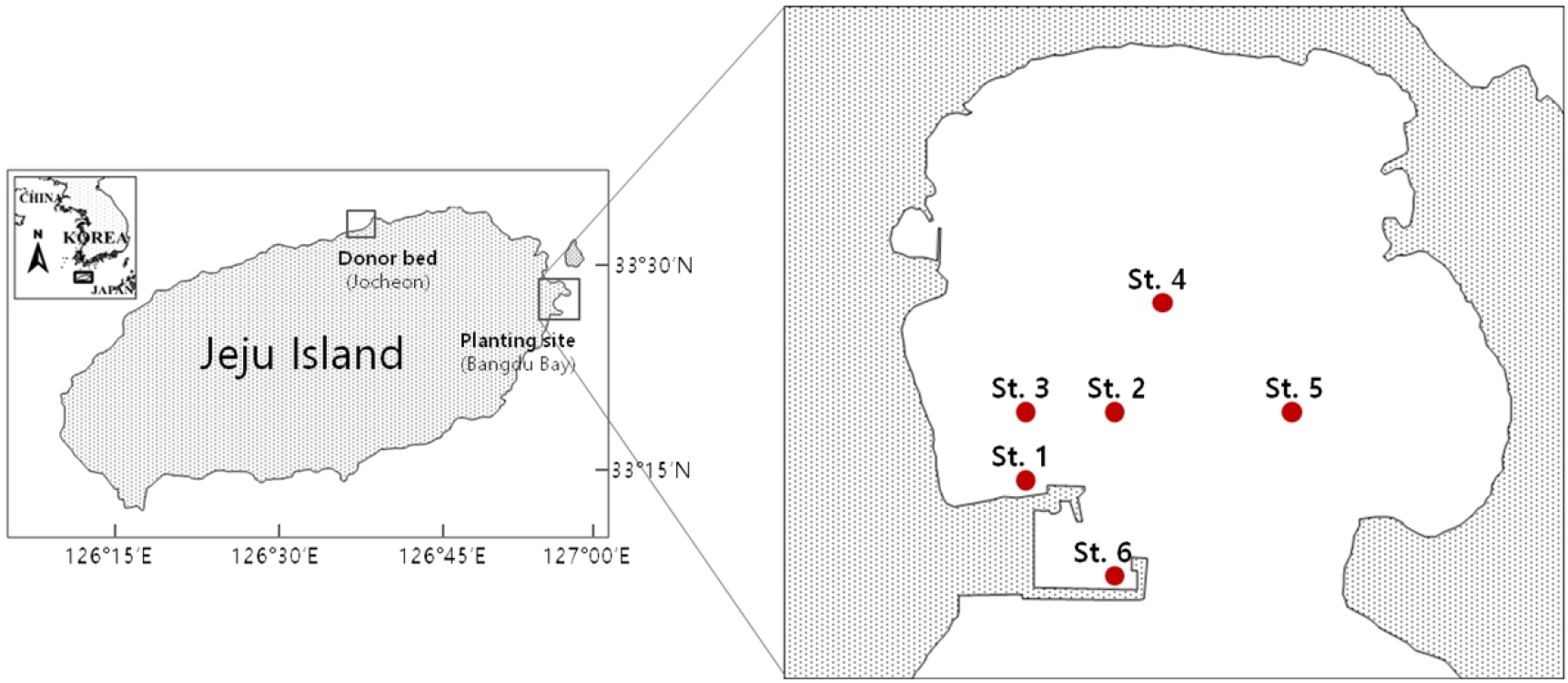
Table 1.
Location of Zostera marina transplanted sites in Bangdu Bay
2.2 생육 환경 조사
수중광량과 수온을 조사하기 위해 각 이식 장소 중심부의 거머리말 서식 수심에 센서(HOBO onset Pendant Temperature/ Light Logger, Onset Computer, USA)를 설치하여 15분마다 조도와 수온을 측정하였다. 측정된 조도(lumens ․ ft-2)는 동 시간에 광량 측정기(LI-1400, LI-COR, Inc)로 측정된 수치와의 회귀분석을 통하여 수중광량(PFD, photon flux density)으로 변환하였다(Lee et al., 2005; Li et al., 2010). 각 이식 장소의 수중광량(mol photons m-2․ d-1)과 수온(°C)은 각각 월평균 값으로 나타내었다. 염분 농도는 2020년 거머리말 이식이 이루어진 St. 3 ~ St. 6의 각 이식 장소 중심부의 거머리말 서식 수심에 자동 전도도 측정 저장 센서(Odyssey Conductivity/Temperature Logger)를 설치하여 15분마다 전도도를 측정하여 월평균 염분 농도(psu)로 계산하였다. 부유성 해조류인 파래의 조사는 매 조사 시 각 장소의 거머리말 군락지에 누적되는 파래의 높이(cm)를 측정하여(n=4) 평균값으로 나타내었다. 조사 지역을 관통한 태풍은 2019년 9월 6일 13호 “링링”과 9월22일 17호 “타파”, 2020년 8월 25일 8호 “바비”, 9월 2일 9호 “마이삭”과 9월 7일 10호 “하이선”으로 조사지역을 관통할 당시의 태풍 강도는 기상청 자료(http://www.weather.go.kr/weather/typoon/typhoon_06_01.jsp)를 활용하였다(Table 2).
Table 2.
Typhoons passed through the study sites during the survey period
2.3 이식 거머리말 조사
이식 거머리말(transplants)의 밀도는 매월 각 이식 장소의 동일한 지점에 방형구(25 cm × 25 cm) 내에 서식하는 모든 영양지(vegetative shoot)와 화지(flowering shoot)의 개체수를 측정하였으며(n=4), 단위면적당 생육밀도(shoots m-2)로 나타내었다. 거머리말의 초기 생존율은 각 이식 장소에서 영양번식으로 측지가 형성되어 밀도가 증가하기 전의 최저 개체수를 측정하여(n=4) 백분율(%)로 나타내었다(Park and Lee, 2007). 이식 거머리말의 밀도 증가율은 철사고정법으로 이식한 거머리말의 일반적인 착생기간으로 보고되는(Park et al., 2009; Li et al., 2010) 3개월 후와 이식지에서 4계절이 경과한 10개월 후, 각 이식 장소에 생존한 개체수를 측정하여(n=4) 백분율(%)로 나타내었다.
거머리말의 형태적 특성 조사를 위해 매 조사 시 잘피의 잎 수, 잎 폭, 잎 길이, 엽초 폭, 엽초 길이와 개체 길이를 수중에서 측정하여(n=6~15) 평균값으로 나타내었다. 잎 수는 엽초 위로 출현한 모든 잎의 수를 계수하고, 잎 폭은 가장 긴 잎의 중간 부분의 폭(mm), 잎 길이는 엽초 상단부터 잎 선단까지의 길이(cm)를 측정하였다. 엽초 폭은 엽초의 가장 넓은 부분의 폭(mm), 엽초 길이는 엽초 하단에서 상단의 길이(cm)를 측정하였고, 개체 길이(cm)는 잎 길이와 엽초 길이의 합으로 구하였다(Park et al., 2009; Li et al., 2010).
2.4 통계 분석
통계 분석은 SPSS 10.1을 이용하였으며, 모든 측정치는 평균(mean)과 표준오차(SE)로 나타내었다. 통계 분석 전 모든 자료의 normality와 homogeneity of variance를 검정하였고, 분석값이 유의할 경우, 각각 Turkey HSD (Honestly Significant Difference)와 Duncan의 사후검증을 시행하였다. 환경 요인 즉, 수중광량, 수온, 염분, 누적 파래의 높이와 이식 잘피의 밀도, 형태적 특성과 밀도 증가율은 two-way ANOVA를 이용하였고, 각 장소의 생존율은 one-way ANOVA를 이용하여 분석하였다. 각 장소의 환경 요인과 이식 잘피의 생육밀도, 잎 폭과 개체 길이의 상관성은 피어슨 상관관계 분석을 이용하여 평가하였다.
3. 결 과
3.1 생육 환경
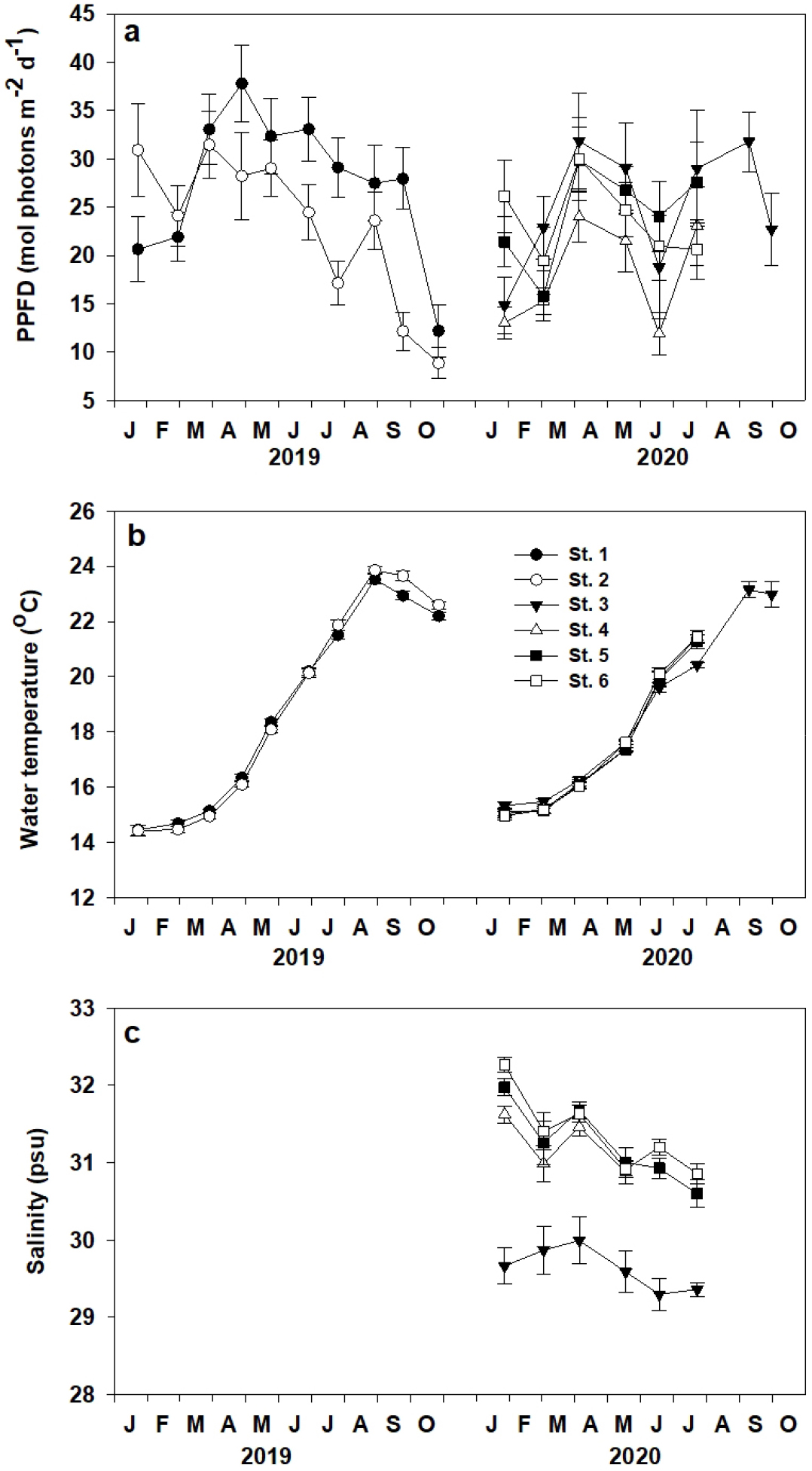
이식 장소의 수중광량은 조사시기마다 유의한 차이(p<0.001)가 나타났으며, 모든 장소에서 봄에 최대값을 보였고 가을 이후 급격히 감소하였다(Fig. 2a). 조사기간 평균 수중광량은 23.6±6.5 mol photon m-2d-1이었으며, St. 1에서 27.6±2.4 mol photon m-2d-1로 가장 높았고, St. 4에서 18.1±2.2 mol photon m-2d-1로 가장 낮았다. 수온도 조사시기마다 유의한 차이(p<0.001)가 나타났으며, 조사장소별로도 유의한 차이(p<0.01)를 보였다(Fig. 2b). 조사기간 평균 수온은 18.2±4.6°C이었고, 모든 장소에서 이식 후 증가하여 여름에 최대를 보이는 계절경향이 나타났다. 월평균 수온은 St. 2에서 2019년 1월 14.4±0.2°C로 최저값과 2019년 8월 23.9±0.1°C로 최대값이 나타났다. 각 장소의 염분도 조사시기마다 유의한 차이(p<0.001)가 나타났으며, 조사장소별로도 유의한 차이(p<0.001)를 보였다(Fig. 2c). 조사기간 평균 염분은 30.9±7.5 psu이었으며, St. 3에서 29.6±0.1 psu로 가장 낮았고, St. 6에서 31.4±0.2 psu로 가장 높았다.
파래 누적 높이는 조사 장소별 시기별 유의한 차이(p<0.001)를 나타내었다(Fig. 3). 각 장소의 파래 출현은 St. 1과 St. 2에서는 2019년은 5월, St. 3, St. 4와 St. 5에서는 2020년 3월, St. 6에서는 2020년 4월부터 시작되었다. St. 3 ~ St. 6의 파래 누적 높이는 수온의 증가와 함께 높아지는 경향을 보였으며(Table 3), 조사장소별 파래의 최대 누적 높이는 2020년 6월 St. 3에서 36.3±5.1 cm로 가장 높았고, 2019년 6월 St. 1에서 8.0±0.8 cm로 가장 낮았다.
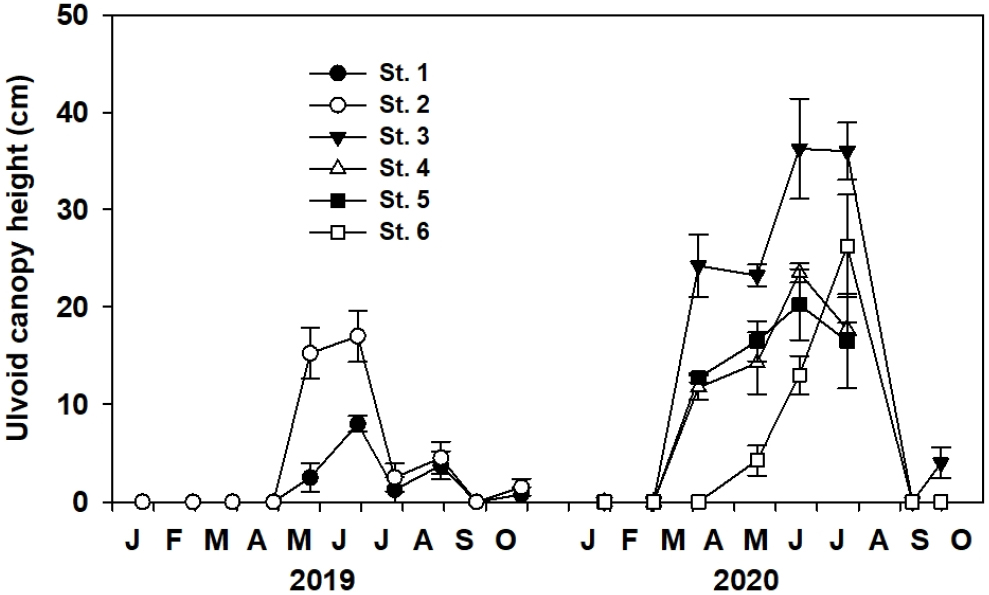
Table 3.
Pearson's correlation analysis results (two-tailed) for underwater irradiance (UI), water temperature (WT), Salinity (SAL), Ulva canopy height (UL), shoot density (SD), leaf width (LW), shoot height (SH)
| UI | WT | SAL | UL | SD | LW | SH | ||||||||||||||||||
| St. 1 | UI | - | 0.747 | ND | - | 0.077 | - | 0.341 | - | 0.524 | - | 0.499 | UI | St. 2 | ||||||||||
| WT | - | 0.386 | ND | 0.599 | 0.450 | 0.675 | 0.601 | WT | ||||||||||||||||
| SAL | ND | ND | ND | ND | ND | ND | SAL | |||||||||||||||||
| UL | 0.295 | 0.642 | ND | - | 0.076 | 0.274 | 0.218 | UL | ||||||||||||||||
| SD | 0.720 | 0.891 | * | ND | 0.545 | 0.561 | 0.270 | SD | ||||||||||||||||
| LW | 0.475 | 0.617 | ND | 0.174 | 0.555 | 0.889 | ** | LW | ||||||||||||||||
| SH | 0.091 | 0.652 | ND | 0.103 | 0.455 | 0.715 | SH | |||||||||||||||||
| St. 3 | UI | 0.216 | - | 0.147 | 0.232 | 0.678 | - | 0.838 | * | - | 0.858 | UI | St. 4 | |||||||||||
| WT | 0.241 | - | 0.534 | 0.861 | * | 0.570 | - | 0.606 | - | 0.456 | WT | |||||||||||||
| SAL | 0.253 | 0.033 | - | 0.257 | - | 0.163 | 0.158 | 0.194 | SAL | |||||||||||||||
| UL | 0.458 | 0.905 | * | - | 0.638 | 0.801 | - | 0.573 | - | 0.489 | UL | |||||||||||||
| SD | 0.642 | 0.744 | - | 0.484 | 0.889 | * | - | 0.840 | * | - | 0.686 | SD | ||||||||||||
| LW | - | 0.279 | 0.552 | - | 0.577 | 0.187 | - | 0.048 | 0.707 | LW | ||||||||||||||
| SH | - | 0.138 | 0.853 | ** | - | 0.974 | 0.688 | 0.585 | 0.408 | SH | ||||||||||||||
| St. 5 | UI | - | 0.461 | 0.472 | - | 0.516 | 0.127 | - | 0.332 | - | 0.220 | UI | St. 6 | |||||||||||
| WT | 0.468 | - | 0.719 | 0.949 | ** | 0.705 | 0.128 | 0.478 | WT | |||||||||||||||
| SAL | - | 0.200 | - | 0.857 | * | - | 0.615 | - | 0.804 | - | 0.355 | - | 0.013 | SAL | ||||||||||
| UL | 0.729 | 0.809 | * | - | 0.697 | 0.485 | 0.110 | 0.357 | UL | |||||||||||||||
| SD | 0.522 | 0.882 | ** | - | 0.869 | * | 0.806 | 0.581 | 0.495 | SD | ||||||||||||||
| LW | - | 0.524 | - | 0.025 | - | 0.079 | 0.044 | - | 0.121 | 0.445 | LW | |||||||||||||
| SH | - | 0.561 | - | 0.183 | 0.267 | - | 0.407 | - | 0.178 | 0.536 | SH | |||||||||||||
3.2 이식 거머리말
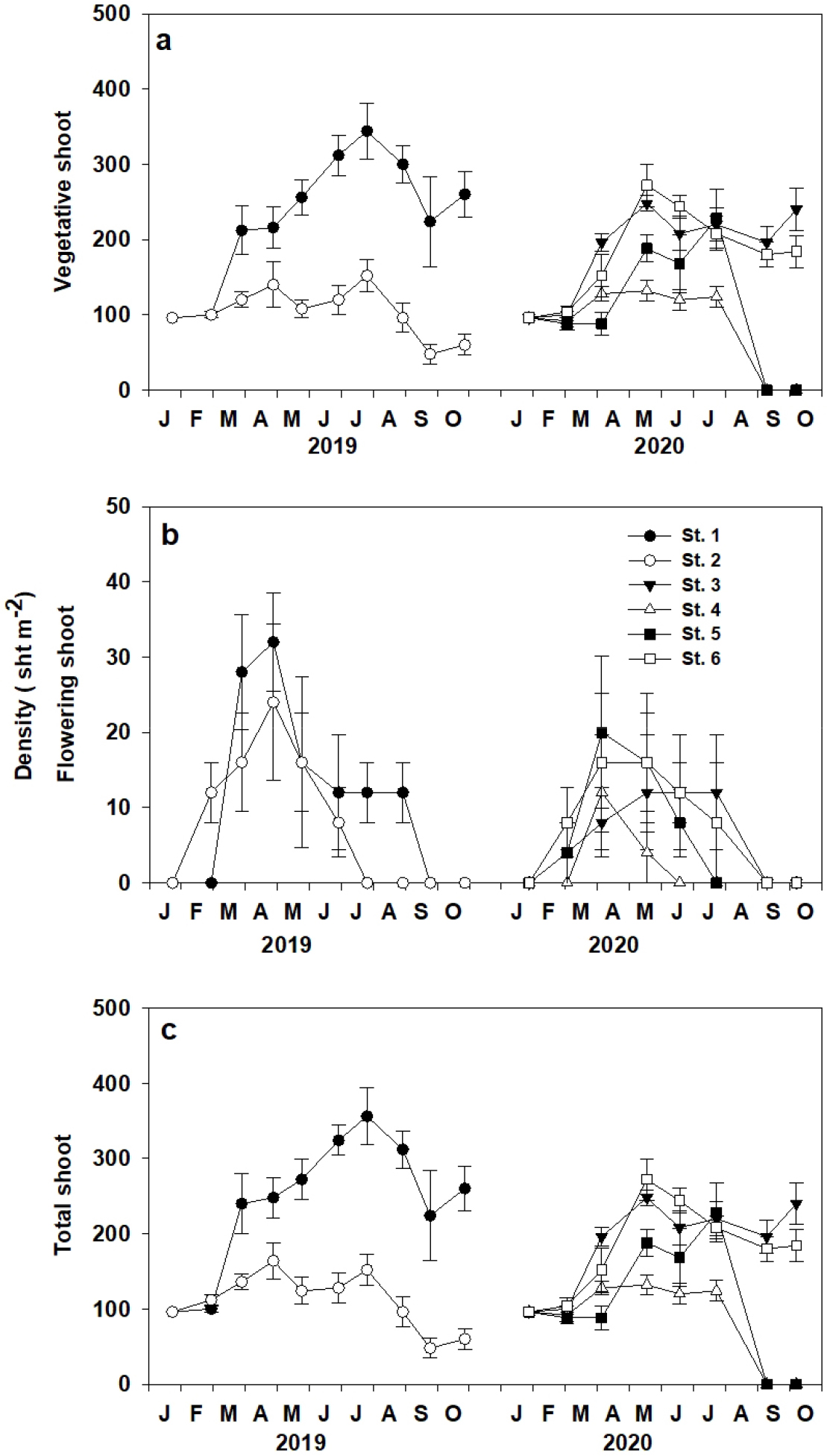
이식 거머리말의 생육밀도는 조사 장소별 시기별 유의한 차이(p<0.001)가 나타났다(Fig. 4). 모든 이식지에서 영양지의 밀도는 이식 직후부터 봄과 초여름 동안 급격히 증가하였으며, 최대 영양지 밀도는 St. 1에서 2019년 7월 344.0±37.2 sht m-2로 가장 높았고, St.4에서 2020년 4월 132.0±13.7 sht m-2로 가장 낮았다(Fig. 4a). 화지는 모든 조사장소에서 봄에 출현하여 여름에 탈락하였으며, 최대 화지 밀도는 St. 1에서 2019년 4월 32.0±6.5 sht m-2로 가장 높았고, St. 3과 St. 4에서 2020년 3월과 4월 12.0±7.7 sht m-2로 가장 낮았다(Fig. 4b). 이식 거머리말의 총밀도도 봄에 급격히 증가한 후 여름 이후 감소하다가 가을에 조금 증가하는 경향이 나타났다. 조사 장소별 최대 서식밀도는 St. 1에서 2019년 7월 356.0.0±38.3 sht m-2로 가장 높았고, St.4에서 2020년 4월 140.0±13.7 sht m-2로 가장 낮았다(Fig. 4c).
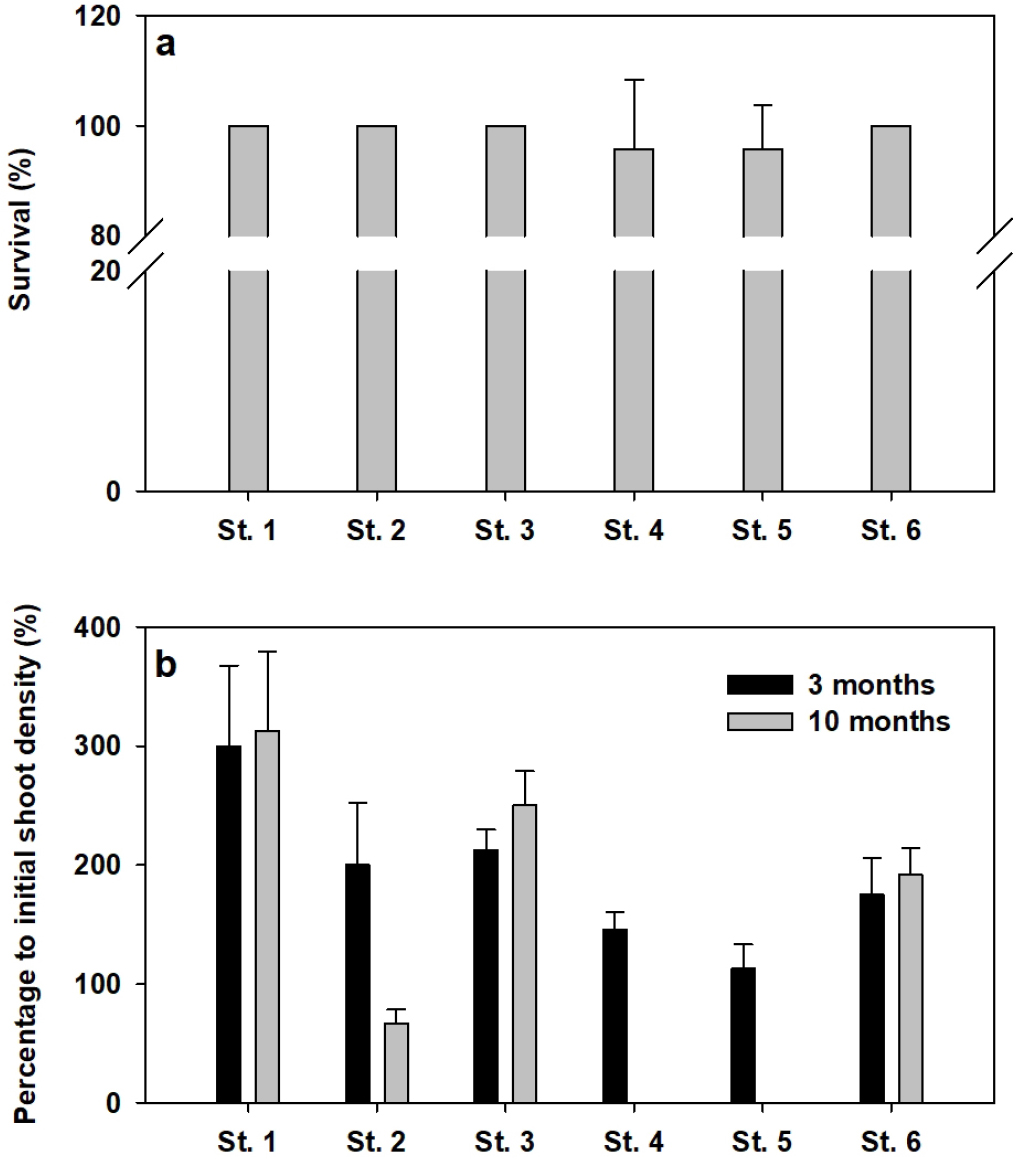
이식 거머리말의 생존율은 조사장소에 따라 유의한 차이가 나타나지 않았으며(p=0.984), 모든 장소에서 대부분 이식 개체가 초기 유실 없이 전량 생존하였다(Fig. 5a). 이식 거머리말의 3개월과 10개월 후의 밀도 증가율은 조사장소에 따라 유의한 차이(p<0.001)가 나타났다(Fig. 5b). 이식 잘피의 3개월 후 평균 밀도 증가율은 191.0±26.4%로, St. 1에서 300.0±67.7%로 가장 높았고 St. 5에서 112.5±20.8%로 가장 낮았다. 10개월 후의 평균 밀도 증가율은 205.2±42.7%이었고, St. 1에서 312.5±66.8%로 가장 높았으나 St. 4와 St. 5에서는 이식 잘피가 2020년 8월 태풍의 영향으로 모두 유실되었다.
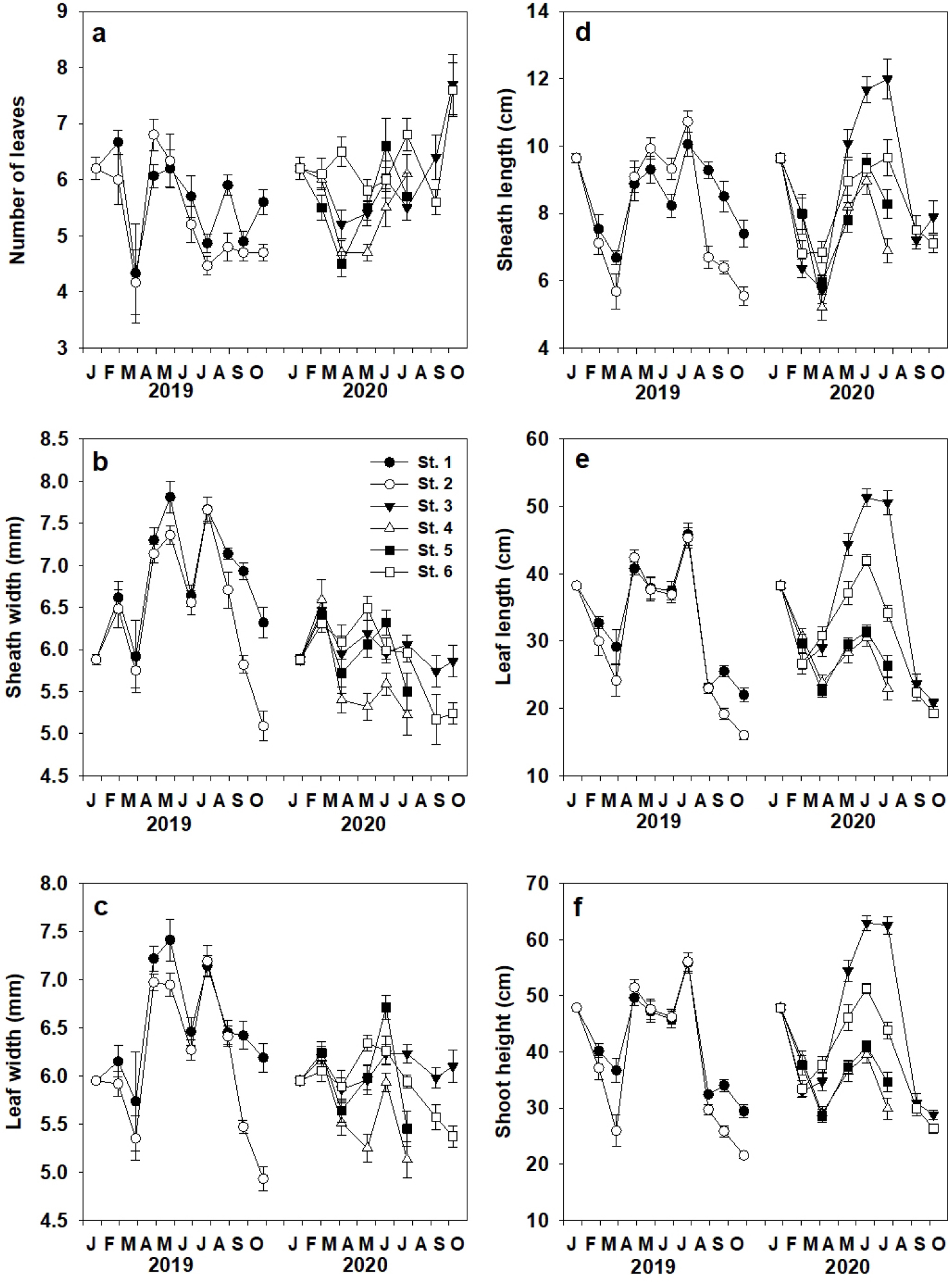
이식 잘피의 형태적 특성 즉, 잎 수, 엽초 폭과 잎 폭, 엽초 길이, 잎 길이와 개체 길이는 조사 장소별 시기별 유의한 차이(p<0.001)가 나타났다(Fig. 6). 이식 잘피의 평균 잎 수는 5.8±0.1개 이었고, St. 6에서 6.3±0.2개로 가장 많았고, St. 2에서 5.3±0.3개로 가장 작았다(Fig. 6a). 잘피 개체의 평균 엽초 폭과 잎 폭은 각각 6.1±0.2 mm과 6.0±0.1 mm로, St. 4를 제외한 모든 장소에서 이식 후 2개월이 경과하면서 증가하였다. 평균 엽초 폭과 잎 폭은 St. 1에서 각각 6.8±0.2 mm와 6.5±0.2 mm로 가장 넓었고, St. 4에서 각각 5.7±0.2 mm와 5.7±0.2 mm로 가장 좁았다(Fig. 6b, c). 이식 잘피의 평균 엽초 길이와 잎 길이는 각각 8.3±0.1 cm와 31.7±1.0 cm로, 이식 2개월까지 감소한 후 증가하였다. 평균 엽초 길이와 잎 길이는 St. 3에서 각각 8.8±0.8 cm와 35.6±4.3 cm로 가장 길었고, St. 4에서 7.8±0.6 cm와 29.1±2.3 cm로 가장 짧았다(Fig. 6d, e). 잘피의 평균 개체 길이는 39.9±1.1 cm이었고, 이식 후 2개월까지 감소한 후 증가하였으며, St. 3에서 44.4±5.1 cm로 가장 길었고, St. 4에서 36.9±2.8 cm로 가장 짧았다(Fig. 6f).
4. 토 론
이식된 잘피의 초기 생존율 확보는 잘피 이식의 성공 여부를 좌우하는 중요한 인자이다(Davis and Short, 1997; Paling et al., 2001). 잘피 이식에서 초기 생존율이 확보되지 못하는 대부분의 경우는 이식된 잘피, 특히 지하부가 이식지에 정착하지 못하고 유실된 결과 발생한다. 이식된 잘피는 이식지에서 새로운 지하부 조직 즉, 지하경과 뿌리가 자라나 퇴적물에 정착해야 새로운 측지를 형성하여 밀도를 증가시킨다(Park and Lee, 2007; Park et al., 2009, 2011). 양성부력을 가진 잘피는 이식지의 퇴적 환경과 조류의 세기 등을 감안한 안전한 식재 방법으로 이식하여야 유실을 방지하여 초기 생존율을 확보할 수 있다. 본 연구 결과 방두만내 6개의 장소에 이식된 거머리말 개체들은 대부분 이식 초기 밀도 감소 없이 높은 생존율을 보이며 정착하여 모든 장소에서 밀도 증가가 나타났다. 이는 본 이식 실험에서 사용한 거머리말의 지하경을 철사로 꽂아 직접 식재하는 철사고정법이 방두만에 적합한 거머리말 이식방법임을 알 수 있다.
이식지의 수중광량, 수온과 염분 농도는 이식된 잘피의 생존율과 생장을 제한하는 중요한 요소이다. 우리나라 연안에서 건강한 거머리말 군락지에서의 평균 수중광량은 8.6 mol photons m-2d-1 이상으로 보고되며(Lee et al., 2005; Park et al., 2009, 2011), 일정기간 이상 5 mol photons m-2d-1 이하의 광이 공급되면 거머리말은 원활한 광합성이 이루어지지 못해 타격을 입게 된다(Zimmerman et al., 1995; Park et al., 2013; Eriander, 2017). 방두만의 거머리말 이식 실험 장소 6개소의 조사기간 중 평균 수중광량은 23.6 mol photons m-2d-1이었고, 특히 이식 후 정착과 생장이 가장 활발한 5개월간의 평균 수중광량은 24.9 mol photons m-2d-1로 조사되었다. 또한, 각 장소의 최저 월평균 수중광량은 13.8 mol photons m-2d-1이었고, 최소값이 나타난 St. 2에서도 2019년 10월 8.9 mol photons m-2d-1로 우리나라 평균 거머리말 군락지의 수중광량보다 조금 높게 나타났다. 즉, 방두만 6개 조사 장소에는 거머리말이 원활하게 생장할 수 있는 충분한 수중광량이 제공된 것으로 보였으며, 이곳의 수중광량은 이식 거머리말의 밀도와 형태적 특성에 유의한 상관관계가 나타나지 않았다(Table 3).
우리나라 연안의 거머리말은 봄부터 초여름 동안 수온 범위가 15-20°C에서 가장 생장이 활발하고, 여름 수온이 30°C 이상에 노출되면 거머리말의 생장과 생체량의 급격한 감소가 일어난다(Lee et al., 2005, 2007). 이는 고수온에서 거머리말의 호흡률의 증가로 광합성율이 감소하기 때문으로 안정적인 거머리말 서식지 조성을 위한 후보 장소 선정에서도 여름 수온이 28°C 미만의 해역이 권장된다(FIRA, 2019). 방두만은 약 64 ha의 좁은 만이나 얕은 연안에서 난류에 의해 저층 흐름이 급격히 변하여(Min et al., 2020) 각 이식 장소의 관측수온도 유의한 차이가 나타났을 것으로 추측되나, 방두만 내 각 이식 장소의 수온은 이식 거머리말의 밀도나 형태적 특성과 유의한 상관관계가 나타나지 않았다(Table 3). 방두만내 모든 조사장소의 조사기간 중 평균 수온은 18.2°C로 거머리말의 생장이 활발한 온도 범위였고, 여름 최대 수온이 22.4°C로 잘피 생육에 해를 끼칠 수 있는 고수온에 노출되지 않는 안전한 범위에 속한다고 볼 수 있다.
해수에 적응되어 살아가는 잘피의 생장과 생존은 염분의 변화에 민감하게 반응할 수 있다(Fernández-Torquemada and Sánchez-Lizaso, 2005). 하구나 만에 위치한 잘피서식지는 일시적으로 저염분수(2-5 psu)나 고염분수(35-40 psu)에 노출되기도 하지만(Koch et al., 2007), 과다한 담수의 지속적 유입으로 저염분 상태가 장기화되어 서식지가 훼손된 사례도 있다(Park et al., 2011). 방두만 북서쪽에는 해저 지하수가 일평균 170 cm d-1의 속도로 용출되지만(Hwang and Koh, 2012), 조사장소 잘피군락지의 잎 위치에서 측정된 월평균 염분 농도는 30.9 psu로 전형적인 해양성 염분농도로 분석되었다. St. 3에서의 평균 염분농도는 29.6 psu로 비교적 낮았으나, 잘피의 생존율이나 밀도 증가율은 가장 높게 나타났다. 각 장소의 염분 농도와 거머리말 서식 밀도는 상관관계에서 대부분의 조사장소에서 유의한 상관관계가 나타나지 않아 방두만 내 염분 농도는 이식 거머리말의 생존율에 결정적인 영향을 미치지 않는 것으로 보인다.
잘피의 이식 적지 판정 기준에서 경쟁생물의 유무나 그 개체수는 매우 중요한 요소이다. 이식 잘피의 생장을 방해하는 잘피 엽상체를 피복하는 경쟁식물의 밀도나 잘피를 섭식하는 민꽃게류 등 조식 동물의 포식압이 과도하지 않아야 한다(Nelson and Lee, 2001; Sugimoto et al., 2007; FIRA, 2019). 방두만 해안은 2000년대 이후부터 과도하게 발생되는 구멍갈파래가 해변으로 밀려와 심각한 환경문제를 초래하였다(Song et al., 2016). 구멍갈파래는 해수중의 무기영양염류를 빠르게 흡수하여 짧은 기간에 급속한 생장율을 보여 다른 해양식물의 생육공간을 선점하게 된다(Nelson and Lee, 2001). 정체된 수역에서 구멍갈파래가 두터운 매트를 형성하여 잘피를 뒤덮게 되면 잘피는 충분한 수중광량을 공급받지 못해 광합성 저해로 생장이 둔화되거나 사멸하게 된다. 태화강 하구에 조성된 잘피군락지에서도 급속한 갈파래의 생장으로 잘피 잎의 60%가 뒤덮힌 일부 군락지는 대부분의 잘피가 사멸하였다(Park et al., 2013). 방두만에서도 구멍갈파래는 모든 장소에서 봄부터 출현하여 초여름에 최대 높이를 나타내었다. 구멍갈파래 축적이 비교적 심했던 St. 3, St. 4와 St. 5의 구멍갈파래의 축적 높이는 2020년 6월 각각 이식된 거머리말 잎의 47.9%, 47.7%와 34.1%에 해당하는 높이를 덮었다. 그러나 이 시기 각 장소의 월 평균 수중광량은 각각 18.8 mol photons m-2d-1, 11.9 mol photons m-2d-1과 24.0 mol photons m-2d-1로 높았고, 수중광량과 파래 높이는 유의한 상관관계가 나타나지 않았다(Table 3). 또한, 대부분의 장소에서 축적된 파래 높이와 거머리말 서식 밀도도 유의한 관계가 나타나지 않았으며, 파래 축적이 가장 높았던 St. 3에서 오히려 약한 양의 상관관계가 나타났다(Table 3). 이는 조사장소의 파래는 좁은 만내에서 빠른 저층 조류의 흐름으로 거머리말 군락에 지속적으로 정체되지 않고 해수 중에서 조류에 따라 움직이거나 해안으로 밀려나가 이식 거머리말의 생존과 생장을 저해하는 요인으로 영향을 미치지 않는 것으로 유추된다.
이식용 잘피는 채취 과정에서 지하경, 뿌리나 잎 조직이 손상되거나 운반, 이식 과정 중 공기 중에 노출되기도 하여 불가피한 이식 충격에 노출된다. 이식 충격은 일반적으로 잘피의 형태 감소로 나타나게 되고 심한 경우 이식 개체의 생존율 감소로 이어지기도 한다(Meinesz et al., 1993). 일반적인 이식 충격은 일정 기간 잘피가 이식지에 적응하면서 회복되고, 이 후 잘피는 형태나 밀도의 증가가 나타난다. 우리나라 연안에서도 거머리말 이식 후 1~3개월 동안 잎 폭과 길이, 개체 길이 등의 형태가 감소하다가 형태와 밀도가 증가한 거머리말 이식 결과가 보고되기도 하였다(Park et al., 2009, 2011; Li et al., 2010, 2013). 방두만의 6개소에 이식된 거머리말도 이식 초기 밀도 감소가 없이 대부분 1~2 개월 간 잎 수, 폭과 길이, 엽초 폭과 길이 감소가 나타난 후 증가하는 것이 관찰되었다.
대형 태풍은 위협적인 풍속으로 인한 퇴적물의 침식, 이동뿐만 아니라 동반되는 강우 등으로 인하여 잘피군락지에 피해를 입히지만(Kim et al., 2015), 잘피군락지에 피해를 입히지 않는 경우 또한 보고된 바 있다(Anton et al., 2009). 조사 기간 동안 이식 후 8~9개월이 경과한 시기에 총 5회의 대형 태풍 즉, 2019년 9월에 태풍 13호 “링링”과 17호 “타파”, 2020년 8월말부터 9월 초 동안 8호 “바비”, 9호 “마이삭”과 10호 “하이선”이 관통하였다(Table 2). 우리나라 연안의 잘피는 봄과 초여름 동안 높은 밀도 증가율을 보이며 서식 밀도가 급격히 증가하고, 늦여름 이후 서식 밀도가 감소하는 계절변화를 보인다(Lee et al., 2005, 2007; Park et al., 2009). 방두만 서쪽에 위치한 실험 장소인 St. 1, St. 2, St. 3과 St. 6의 거머리말 서식 밀도는 이러한 일반적인 경향이 나타났지만, 만의 중심부에 위치한 St. 4, St. 5의 이식 거머리말은 태풍 이후 모두 소실되었다. 이는 조사 지역을 관통한 대형 태풍의 영향이 만의 중심부에 위치한 이 2개소에 더 강하게 미친 결과로 유추된다.
결론적으로 방두만의 6개 후보 장소에 이식된 거머리말은 모든 장소에서 대부분 생존하였고, 착생 후 높은 밀도 증가율을 보여 방두만의 거머리말 군락 조성이 가능함을 확인하였다. 그러나 빈번히 태풍이 통과하는 조사장소의 지협적인 특이성을 감안하면 대형 태풍 후에도 높은 밀도 증가율을 보인 만의 서쪽 연안이 방두만 거머리말 서식지 복원을 위한 이식 적지로 판단된다.
