

1. 서 론
저서동물은 퇴적물 내의 유기물을 표면으로 운반해주고, 서식 활동으로 인해 퇴적물 내의 영양분을 수층으로 부유시켜 다른 동물들의 먹이가 되게 하는 등 생태계 내의 연결자 역할을 한다(Diaz et al., 2004; Dauvin et al., 2007). 특히 저서동물은 환경변화에 민감하며, 이동성이 적고, 번식력이 매우 강하여 주변 환경 변화를 연구하는데 장기적인 지표로 사용할 수 있다. 그러나 다양한 분류군이 포함된 저서동물을 이용하여 해양환경 변화를 파악하는데 많은 비용과 시간이 소요된다. 이에 따라 저서동물의 주요 분류군을 이용한 저서생태계 평가가 제안되었다(Jung et al., 2011). 일반적으로 저서다모류는 해양 연성저질에 서식하는 저서동물 중 종 수, 개체 수 측면에서 가장 우점한 분류군이다. 또한 저서다모류는 해양의 수심(Lee, 1987; Choi and Koh, 1988; Yu et al., 2011), 수온(Long and Lewis, 1987), 퇴적물의 퇴적상(Lim and Choi, 1992; Yu et al., 2011), 퇴적물 내의 유기물 함량(Lim et al., 1992)등 다양한 환경요인에 영향을 받는 것으로 알려지고 있다. 그러므로 저서다모류의 군집연구는 전체 저서생태계 변화를 빨리 파악하는 데 매우 효과적이다(Sanders, 1958; Rhoads and Young, 1970; Shin et al., 1989).
우리나라 동해안은 해안선을 기준으로 완만한 경사를 보이다 급격히 수심이 깊어지는 양상을 보이며, 대부분의 해안이 사질퇴적상으로 이루어져있다(Kim et al., 2001). 이는 동해안 대부분 연안에서 공통적으로 보이는 특징으로 동해 연안의 저서다모류의 군집 및 분포 특성이 퇴적물의 퇴적상에 의한 차이보다 퇴적상 외의 다른 환경 요인에 더 큰 영향을 받을 것으로 여겨진다. 특히, 동해 연안에 위치한 고리와 울진 지역의 원자력발전소에서는 주변 수온보다 약 7°C~ 9°C 증가된 온배수로 연간 약 44.6~60.8억 톤 배출한다(해양수산부, 2007). 이로 인해 발전소 주변해역의 대형저서동물 군집은 온배수의 영향을 직·간접적으로 받고 있다(Yu et al., 2011; 2013). 동해 연안에 위치한 발전소 주변 해양생태계는 발전소를 기준으로 울진 지역은 북쪽으로 5.5 km, 남쪽으로 9.7 km까지, 고리 지역은 발전소 남동쪽 3.3 km, 남서쪽 5.7 km까지 온배수의 영향을 받는다고 알려지고 있다(Kim et al., 2011; 해양수산부, 2015).
연안에 건설된 많은 발전소들은 해수를 이용하여 발전 장치를 냉각 한 후, 데워진 해수를 바다로 방출하며, 이때 발생하는 온배수는 주변 해양의 수온을 급격히 증가시켜 해양생물에 영향을 끼치며, 해양생태계를 교란시키는 주요 원인으로 알려지고 있다(Gibbons and Sharits, 1974; Durrett and Pearson, 1975; Warwick, 1993; Kim et al., 2007; Teixeira et al., 2009). 이에 따라 발전소에서 방출되는 온배수는 발전소 주변해역 저서생태계를 조절하는 주요 인자로 여겨지고 있다. 최근에 동일한 퇴적상 지역의 온배수가 해양저서동물의 군집 구조에 미치는 환경 영향 연구가 있었으나, 다양한 퇴적 환경의 온배수 해역에서 저서생태계 군집 구조 연구는 매우 제한적이다(Yu et al., 2011; 2013).
그러므로 본 연구는 동해 연안의 다양한 퇴적상을 가진 발전소 취·배수구 주변 해역을 포함한 온배수 영향 해역인 울진과 고리 해역, 온배수 영향이 없는 해역인 후포 해역을 대조구로 조사 지역을 설정하여 저서다모류 종 조성을 통해 군집구조를 파악하고, 다모류 군집구조에 영향을 주는 환경요인을 파악하였다.
2. 재료 및 방법
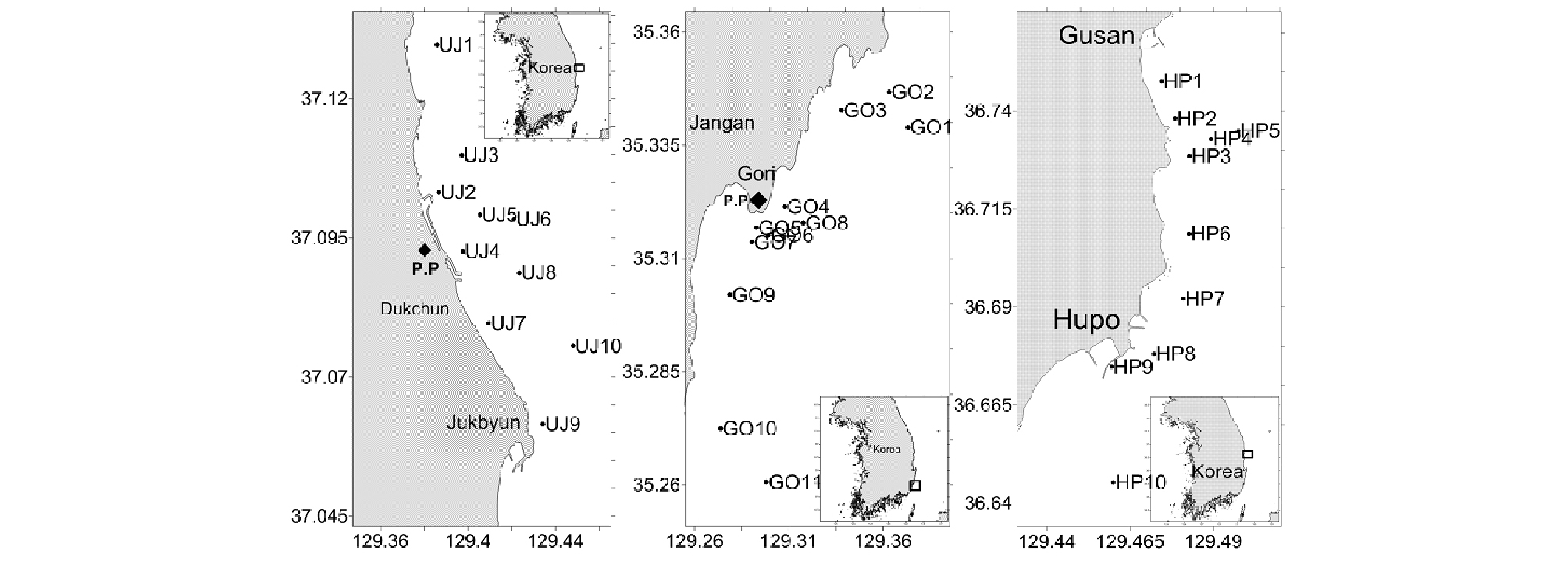
동해 연안의 저서다모류 종 조성 및 군집구조를 파악하기 위하여 2006년부터 2013년까지 울진 10개 정점, 후포 10개 정점, 고리 11개 정점에서 채집을 실시하였다(Fig. 1). 본 연구지역에서 온배수의 영향에 포함된 정점은 1°C 출현 기준 울진은 모든 정점, 고리 7개 정점(ST. 4, 5, 6, 7, 8, 9, 10)이였다(Kim et al., 2011; 해양수산부, 2015). 시료 채집은 Smith-McIntyre 그랩(0.1 m2)을 이용하여 각 정점에서 2회씩 실시하였고, 현장 선상에서 1 mm 네트에 여과 후 생물을 포함한 퇴적물을 10% 중성포르말린으로 고정하였다. 고정된 퇴적물은 실험실로 운반하여 개체수와 습중량을 측정하였고, 현미경을 이용하여 가능한 종 수준까지 동정하였으며, 출현 개체수를 계수하였다. 각 정점으로부터 얻어진 저서다모류의 종별·개체수 자료는 단위면적당(m2)으로 환산하여 분석에 사용되었다.
시료 채집과 병행해 환경 요인 조사를 위해 표층 1 cm 깊이의 퇴적물 온도를 측정하였으며, CTD(SBE-19)를 이용하여 저층수에서 정점별 염분과 수온, 저층 용존산소량(DO)을 측정하였다. 퇴적 환경 요인 분석을 위하여 채집된 표층 퇴적물의 일부를 냉동 보관 후 실험실에서 입도 분석 및 퇴적물 내 총 유기 탄소량(TOC) 등을 분석하였다. 입도 분석은 채취한 퇴적물 시료 중 5 g을 비이커에 담아 10%의 H2O2로 유기물과 0.1 N-HCl로 탄산염을 제거 후, 퇴적물 시료를 4ø체로 조립질과 세립질로 구분하고 4ø이하의 사질 시료는 Ro-tap sieve shaker로 15분간 체질한 후 입도 등급별로 무게 백분율을 구하였으며, 4ø이상의 니질 시료는 2 g에 0.1% calgon 용액을 넣고 교반시킨 후 X-선 자동입도 분석기인 Sedigraph 5000D를 이용, 입도 무게 백분율을 Folk and Ward(1957)의 Inclusive Graphic Method에 의하여 변수를 구하였다. 퇴적물의 총 유기 탄소량은 50°C에서 48시간 건조 후 1 g씩 채취하여 0.1 N 염산으로 탄산염을 제거한 후 CHN 분석기를 통해 구하였다.
분석에 이용된 환경자료는 각 조사 정점에서의 평균값을 이용하였다. 그러나 저층 수온 값은 각 시기별 조사 지역의 기준 수온에 대한 변동 폭(∆T)을 이용하였다. 울진 지역의 기준 수온은 온배수 영향을 받지 않는 최 북측 정점을 기준으로 평균 수심 약 30 m에 위치한 정점 1과 3 그리고 고리 지역은 정점 2와 3의 수온을 기준 수온으로 이용하였으며, 후포 지역은 평균 수심 약 30 m인 정점 3, 4, 5를 기준으로 각 시기별 수온 변동값을 구하였다. ∆T값은 각 정점의 수온과 기준으로 삼은 정점의 평균 수온의 차이로 계산하였다. 저서다모류의 경우, 조사 기간 동안의 평균 값을 이용하여 저서다모류의 정점 별 출현 종수, 개체수, 서식 밀도, 출현 다모류 분류군별 구성 비율, 우점종 등의 분포 특성을 분석하였다. 이들 자료는 Shannon and Wiener (1963)의 종 다양성 지수(Diversity index, H')와 Simpson의 균등도 지수(Evenness, J') 등을 통하여 분석하였고, 다변량 분석을 위해 정점별 종 조성과 그에 따른 그룹별 유사도 분석을 통한 집괴, MDS (non-metric multi-dimensional scaling) 분석을 실시하였다(Clarke and Warwick, 2001). 분석시 일부 높은 개체수로 출현한 종이 유사도 분석에 미치는 영향을 감소시키기 위해 정점별 총 출현 개체수를 4th root transformation하여 Bray-Curtis의 유사도 지수를 사용하였다(Somerfield, 2008). 각 군집의 유사도에 기여한 기여종을 밝히기 위하여 SIMPER (Similarity Percentages Procedure) test를 시행하였으며(Primer 6.0), 저서다모류의 공간 분포에 영향을 주는 환경 요인을 파악하기 위하여 BIO-ENV (Biota-environment matching using step algorithm) test를 실시하였다.
3. 결 과
3.1 환경 요인
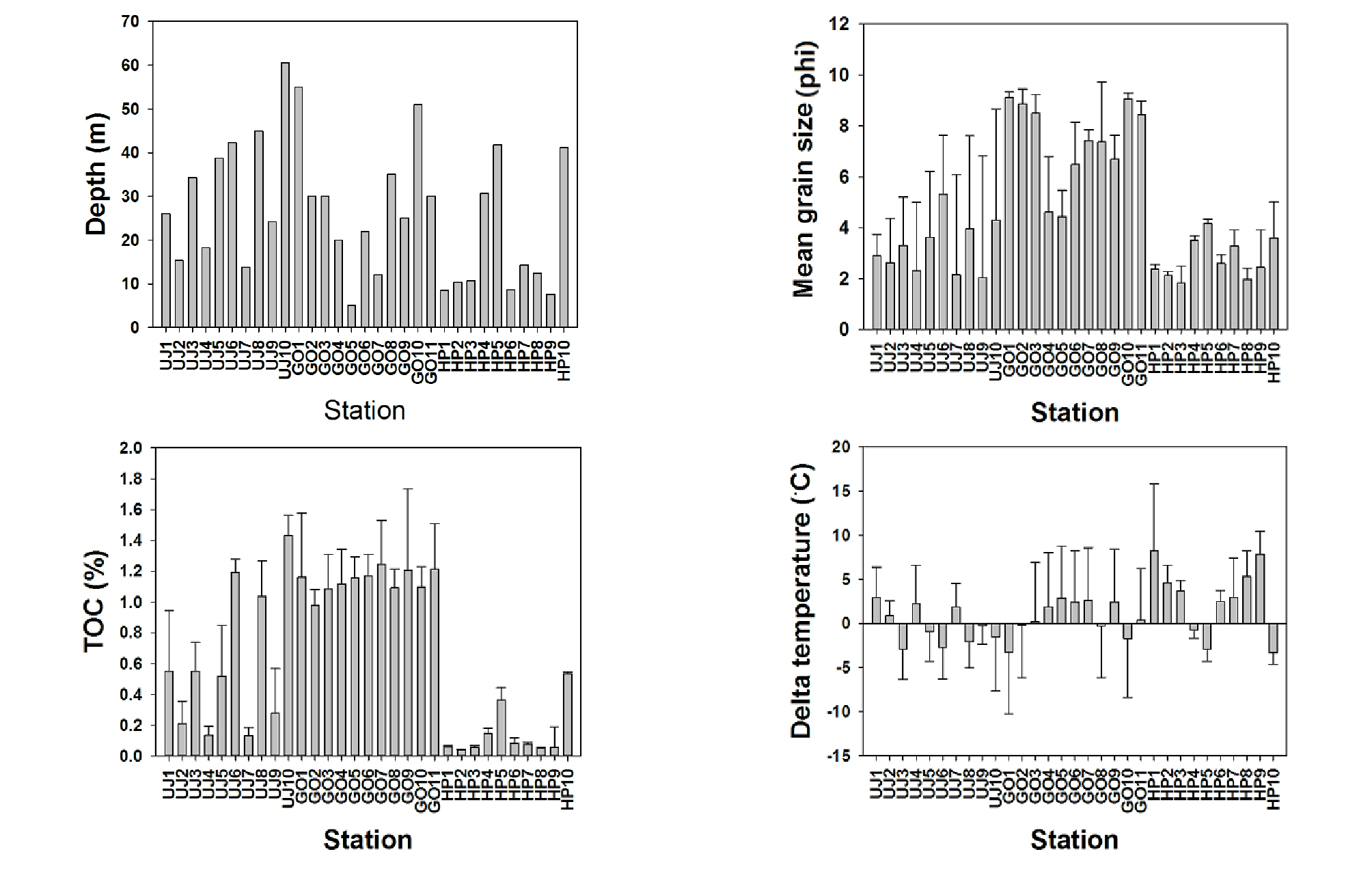
울진과 고리 연안은 발전소가 위치한 지역으로 일부 정점은 온배수의 영향을 받고 있는 지역이며, 후포 연안은 인근의 후포 뱅크가 있어 수심이 비교적 얕고 육지의 영향이 거의 없는 해역이다. 조사 정점의 수심 분포는 약 5~60 m대에 위치하고 있었으며, 평균 수심 26 m에 위치하였다(Fig. 2). 퇴적물의 입도 분포는 1.84~9.11 ø사이였으며, 평균 입도는 4.56 ø로 나타났다. 울진과 후포 연안은 동해 연안의 전형적인 사질 해역으로 대부분의 지역에서 세립한 사질 퇴적상을 보였으나 울진의 6번 정점과 8, 10번 정점은 펄모래 퇴적상을 보였고, 고리 연안은 동해 연안의 대표적인 니질 해역으로 대부분 지역은 니질 퇴적상을 보였으나 4번 정점과 5번 정점은 펄모래 퇴적상을 보였다. 울진과 고리 지역의 취·배수구 주변 정점의 퇴적상은 취·배수구 주변 정점 대부분이 조사기간 동안 크게 변화하는 양상을 보였다. 조사기간 동안의 퇴적물 내 총 유기탄소량은 0.04~1.43% 범위로, 평균 0.65%를 보였다. 총 유기탄소량은 지역별로 유의한 차이를 보였는데, 후포 연안에서 평균 0.15%로 가장 낮았고, 고리 연안은 평균 1.14%로 가장 높았다(Fig. 2). 저층 염분 분포는 전 지역에서 33.3에서 34.7 ppt의 분포를 보였다. 조사 기간 동안 울진 지역 수온 변동(∆T)은 발전소 취·배수구에 인접한 2, 4, 7, 9번 정점에서 -0.2~2.2°C(평균 1.2°C) 사이였다(Fig. 2). 고리 지역의 경우, 취·배수구에 인접한 4, 5, 6, 7, 9번 정점에서 1.9~2.9°C(평균 2.5°C)를 보였으며, 후포 지역은 -3.3~8.2°C(평균 2.8°C) 범위를 보였다. 본 연구 지역은 수심이 깊어질수록 퇴적물이 세립해졌으며, 총 유기탄소량이 증가하고 수온은 감소하는 양상을 보였다.
3.2 종 조성
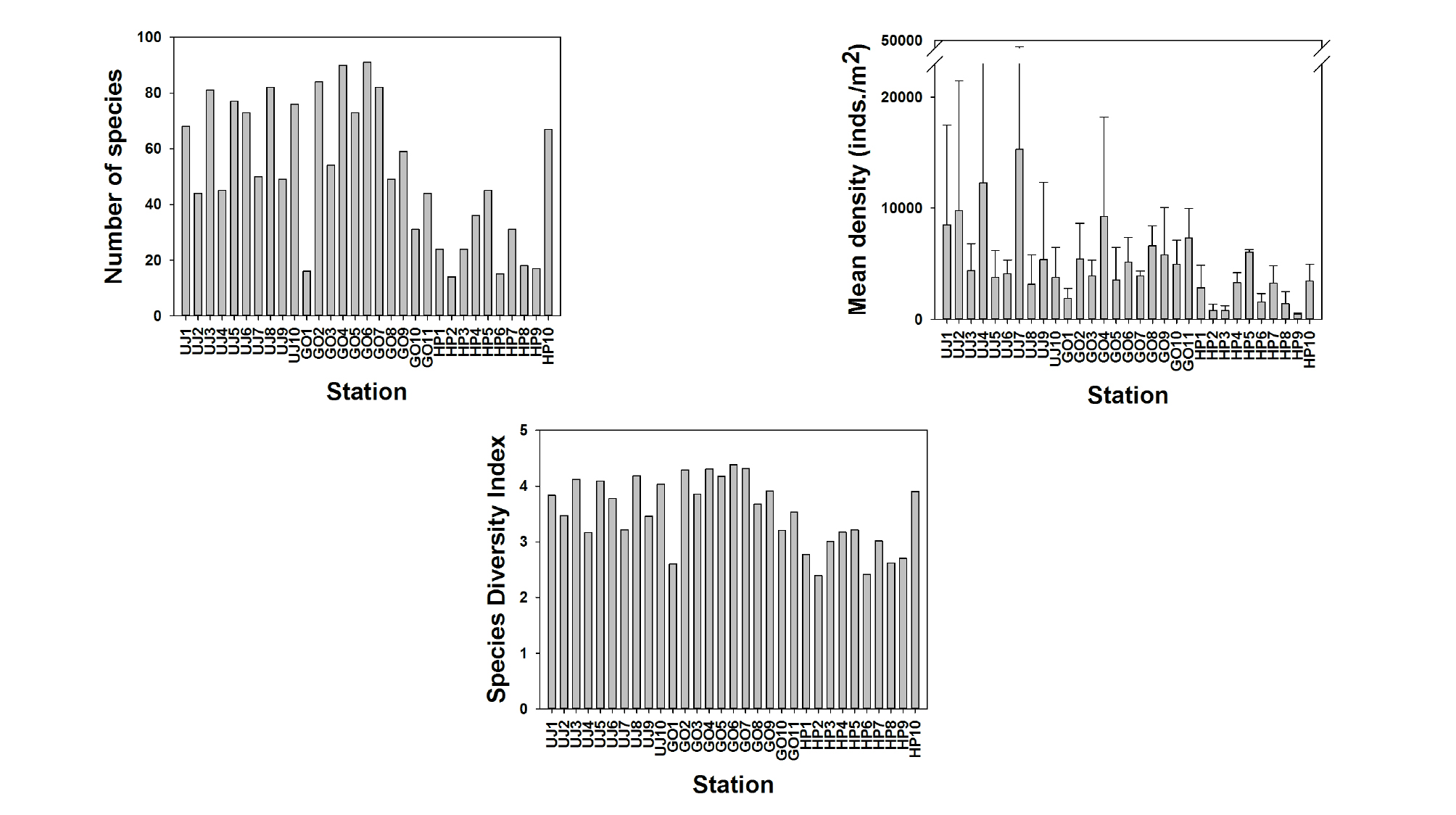
조사 해역에 출현한 저서다모류의 총 종수는 283종이였다. 조사 기간 동안 출현한 저서다모류의 종수는 평균 52종이 출현하였으며, 울진 연안에서 203종으로 가장 많은 종이 출현하였고, 고리 191종, 후포 88종이 출현하였다. 고리의 6번 정점에서 91종으로 가장 많이 출현하였고, 후포의 2번 정점에서 14종으로 가장 적게 출현하였다. 정점별 저서다모류의 출현 개체수는 전체 평균 4,912개체/m2이였고, 울진 해역에서 평균 7,049개체/m2로 가장 많은 개체수가 출현하였으며, 고리 5,253개체/m2, 후포 2,399개체/m2로 가장 적은 개체수가 출현하였다. 울진 연안의 7번 정점에서 15,318개체/m2로 가장 높았으며, 후포 연안의 9번 정점에서 495개체/m2로 가장 낮았다. 조사 기간 동안 각 정점에서 출현한 저서다모류의 종 다양성 지수는 평균 3.51였고, 고리의 6번 정점에서 4.37로 가장 높았으며, 후포의 2번 정점에서 2.39로 가장 낮게 나타났다(Fig. 3). 각 조사지역에서의 종 다양도 지수는 울진 지역에서 평균 3.73, 고리 지역에서 3.84, 후포 지역에서 2.92로 나타났다.
조사 기간 동안 각 정점에서 출현한 우점종은 얼굴갯지렁이과(Family Spionidae)의 민얼굴갯지렁이(Spiophanes bombyx)가 고리를 제외한 지역에서 가장 극우점하는 양상을 보였으며, 송곳갯지렁이과(Family Lumbrineridae)의 긴자락송곳갯지렁이(Lumbrineris longifolia), 얼굴갯지렁이과(Family Spionidae)의 양손갯지렁이(Magelona japonica), 오뚜기갯지렁이과(Family Sternaspidae)의 오뚜기갯지렁이(Sternaspis scutata), 버들갯지렁이과(Family Capitallidae)의 Mediomastus californiensis가 우점하는 양상을 보였다. 조사 기간 동안 우점종의 각 정점별 출현 경향은 최우점종인 민얼굴갯지렁이(S. bombyx)가 울진 지역에서는 5번, 6번, 8번, 10번을 제외한 정점에서 우점 출현하였고, 후포의 전지역과 고리의 4번 정점에서도 많은 수로 출현하였다. 긴자락송곳갯지렁이(L. longifolia)는 울진과 고리 발전소 취·배수구 지역 정점을 제외한 정점에서 주로 출현하였으며, 양손갯지렁이(M. japonica)는 울진과 고리 지역에서는 많은 수로 출현하였으나, 후포 지역에서는 거의 출현하지 않았다. 오뚜기갯지렁이(S. scutata)는 고리의 10번 정점을 제외한 지역에서 많은 수로 우점하였으나, 울진 지역은 매우 적은 수가 출현하였고, 후포 지역은 출현하지 않았다. M. californiensis는 고리 지역에서만 많은 수로 출현하는 양상을 보였다(Fig. 4).

3.3 군집 구조
조사 기간 동안 조사된 저서다모류의 군집 구조는 유사도 20%로 고리 지역(I)과 울진·후포 지역(II)으로 구분되었다(Fig. 5). I 그룹은 퇴적물 내 총 유기탄소량(TOC)이 비교적 높고 퇴적상이 니질이였으며, II 그룹은 TOC값이 비교적 낮고 퇴적상이 사질인 지역으로 나타났다. I 그룹과 II 그룹을 구분하게 하는 종들은 민얼굴갯지렁이(S. bombyx)와 양손갯지렁이(M. japonica), 오뚜기갯지렁이(S. scutata), 긴자락송곳갯지렁이(L. longifolia), M. californiensis 등 대부분 우점종들이 두 그룹간 비유사도에 기여했으며, I 그룹에서는 양손갯지렁이(M. japonica)가, II 그룹은 민얼굴갯지렁이(S. bombyx)가 각 군집을 나누는데 가장 큰 영향을 주는 기여종이였다.(Table 1). 조사 기간 동안 저서다모류의 군집 구조에 영향을 주는 환경요인을 BIO-ENV를 통해 분석하였다. 조사 지역의 저서다모류 군집 구조에 가장 큰 영향을 주는 환경 요인은 퇴적물 내의 총 유기탄소량, 용존 산소량, 퇴적물의 모래 함량, 입도, 퇴적물의 분급도로 나타났으며, 퇴적물 내의 펄 함유량도 군집 구조에 영향을 주는 것으로 나타났다(Table 2).

Table 1. Dominant species contributed to dividing the polychaete community into two distinguished polycheate assemblages in the study area
Table 2. BIO-ENV test to analyze the effect of environmental variables on the polychaete community structure
Table 3. Spearman rank correlation coefficients between the environmental variables and biotic ones from all study sites (*, p<0.05; **, p<0.01; ***, p<0.001; -, p>0.05)
조사 기간 동안 환경요인과 저서다모류의 종 특성과 우점종 간의 상관관계 분석을 실시하였다(Table 3). 저서다모류의 종수와 다양도는 퇴적물의 평균입도와 퇴적물의 총 유기탄소의 양과 유의한 상관관계를 보였으나, 수심과 ∆T와는 유의한 관계를 보이지 않았다(Table 3). 반면에, 저서다모류의 서식밀도는 퇴적물의 총 유기탄소와만 양의 상관관계를 보였다. 주요 우점종들은 퇴적물의 입도와 총 유기탄소의 양이 증가할수록 유의하게 증가하였으나 민얼굴갯지렁이(S. bombyx)는 유의하게 감소하였다. 주요 우점종 가운데, 긴자락송곳갯지렁이(L. longifolia)와 양손갯지렁이(M. japonica)는 ∆T값이 증가할수록 서식 밀도도 같이 증가하는 양상을 보였으나, 나머지 종들은 유의한 관계를 보이지 않았다(Table 3). 온배수의 영향을 받는 두 지역 저서다모류의 종수와 평균 서식밀도, 다양도 및 우점종 들과 환경요인과의 상관관계 분석을 실시한 결과, 사질 지역인 울진 지역의 저서다모류 출현종 수와 다양도가 수심, 퇴적물의 입도, 퇴적물 내 유기탄소량 값, 염분도가 증가할수록 유의한 양의 상관관계를 보였으며, 용존산소량과 퇴적물의 모래 함량이 증가할수록 유의한 음의 상관관계를 보였다(Table 4). 평균 서식밀도는 수심, 퇴적물의 입도, 퇴적물 내 유기탄소량값, 염분도가 증가할수록 음의 상관관계를 가졌으며, 용존산소량과 퇴적물의 모래 함량이 증가할수록 유의한 음의 상관관계를 보였다. 주요 우점종 중 민얼굴갯지렁이(S. bombyx)가 수심, 퇴적물의 입도, 퇴적물 내 유기탄소량값, 염분도가 증가할수록 유의한 양의 상관관계를 가졌으며, 용존산소량과 퇴적물의 모래 함량이 증가할수록 유의한 음의 상관관계를 가진 것으로 나타났다(Table 4). 니질 퇴적상을 가진 고리에서는 저서다모류 출현종 수와 다양도가 수심, 퇴적물의 입도, 염분도가 증가할수록 유의한 음의 상관관계를 보였으며, 용존산소량과 퇴적물의 모래 함량이 증가할수록 유의한 양의 상관관계를 보였다. 평균 서식밀도와 환경요인과는 유의한 상관관계가 없는 것으로 나타났다. 주요 우점종 가운데 긴자락송곳갯지렁이(L. longifolia)는 수심과 퇴적물의 입도, 염분도가 증가할수록 유의한 양의 상관관계를 보였으며, 용존산소량과 퇴적물의 모래 함량이 증가할수록 유의한 음의 상관관계를 보였다. Mediomastus californiensis는 수심과 염분도에 음의 상관관계를 보였고, 용존산소량과 양의 상관관계를 보였다(Table 4).
Table 4. Spearman rank correlation coefficients between the environmental variables and biotic ones from Uljin and Gori coast (*, p<0.05; **, p<0.01; ***, p<0.001; -, p>0.05)
4. 고 찰
우리나라 동해안은 해안선을 기준으로 완만한 경사를 보이다 급격히 수심이 깊어지는 양상을 보인다(Kim et al., 2001). 본 연구의 조사 정점 역시 수심 5~60 m로 해안선에 인접한 위치의 정점에서는 얕고 외해로 향할수록 깊어지는 수심구배를 보였다. 본 연구의 2지역(울진 ․ 후포)은 거의 모든 정점이 사질 퇴적상을 보였으나, 고리 지역은 몇 정점을 제외한 모든 정점이 니질 퇴적상을 보였다. 조사 정점의 퇴적물 내 총 유기탄소량은 수심이 얕고 사질인 정점에서 비교적 낮고 상대적으로 수심이 깊고 니질이 많이 포함된 정점에서 높게 나타나 동해 연안의 일반적인 해양 환경의 특징을 보였다(Paik et al., 2007; Yu et al., 2011).
본 연구 지역에서 출현한 저서다모류의 총 종수와 평균 서식밀도는 각각 283종과 4,912개체/m2이였으며, 울진 연안에서 종수(203종)와 서식밀도(7,049개체/m2)가 높았으며, 고리(191종, 5,253개체/m2)와 후포(88종, 2,399개체/m2) 지역 순으로 감소하였다(Fig. 3). 이는 기존 연구에서 조사된 저서다모류의 종수와 서식밀도보다 높았다. 기존 연구에서 울진 해역의 저서다모류 종수와 평균 서식밀도는 각각 151종, 2,496개체/m2으로 나타났고(Yu et al., 2011), 후포해역(145종과 1,601개체/m2), 고리 해역(101종과 399개체/m2) 순으로 저서다모류의 종수와 서식밀도가 감소하였다(Paik et al., 2007; Kim et al., 2011). 발전소 주변해역인 울진과 고리 지역의 저서다모류는 월성 원자력 발전소 주변 해역에서의 저서다모류 종수(93종) 보다 높았다(Seo et al., 2009). 이는 일반적으로 알려진 채집시기와 채집면적 그리고 종 분석 수준에 따른 차이로 여겨진다. 또한 저서다모류의 종 조성과 서식밀도는 퇴적상의 이질성에 영향을 받는다(Seo et al., 2009). 일반적으로 대형저서동물의 종 조성은 단일 퇴적상 보다는 다양한 퇴적상에서 증가한다(Nishijima et al., 2015). 본 연구 지역에서 조사된 기존 연구에서는 각각의 조사 해역에서의 퇴적상 차이가 크게 나타나지 않았으나, 본 연구에서는 다양한 퇴적상에서 조사되어졌다(Fig. 2). 그러므로 본 연구에서 지역 간 저서다모류의 출현 종과 서식밀도의 차이는 채집시기와 채집면적과 종 분석 수준 그리고 퇴적상의 이질성에 따라 차이를 보이는 것으로 여겨진다. 특히 본 연구에서 출현한 종들을 서식 온도와 관련하여 보면 대부분 광온성에 가깝다. 발전소 주변에선 온수성에 가까운 종들도 나타나지만 전체적으로 볼 때 그 수는 극히 적어 대부분의 개체들이 저층의 낮은 수온에 서식하기에 적합한 환경을 대변하는 것으로 여겨진다.
주요 우점종은 가운데, 울진과 후포지역에서는 민얼굴갯지렁이(S. bombyx), 양손갯지렁이(M. japonica)와 긴자락송곳갯지렁이(L. longifolia)가 우점하였으며, 고리에서는 양손갯지렁이(M. japonica)가 우점하였다(Fig. 4). 본 연구 해역에서의 주요 우점종은 동일 지역에서 조사된 기존 연구와 큰 차이를 보이지 않았다(Paik et al., 2007; Kim et al., 2011; Yu et al., 2011). 민얼굴갯지렁이(S. bombyx)는 퇴적상이 자갈이 거의 없는 세립한 사질 퇴적상 지역에서 우점하였으며, 특정시기에 개체수의 변동 폭이 컸다(Fig. 4). 민얼굴갯지렁이(S. bombyx)는 동해안에서 일반적으로 우점하게 출현하는 종으로 발전소 배수구 주변 자갈의 함량이 매우 낮은 가는 사질 퇴적물에서 주로 출현하며(Yu et al., 2013), 표층 퇴적물에서 먹이를 섭식하는 표층 퇴적물식자로 알려져 있고(Macdonald et al., 2010), 환경 변화에 민감한 기회주의적 종으로 특히 해수의 흐름이 증가하여 퇴적상에 변화가 있을 시 개체수가 급감하는 것으로 알려져 있다(http://www.marlin.ac.uk). 본 조사에서는 민얼굴갯지렁이(S. bombyx)가 울진 발전소 취·배수구 주변 지역에서 극 우점하여 출현하였고, 특히 개체수가 2012년 급증하는 모습을 보였다가 2013년 급감하는 양상을 보였다. 이는 울진 조사 지역의 퇴적상이 2012년 자갈이 거의 없고 세립한 사질 퇴적상에서, 2013년엔 자갈의 함량이 많은 조립한 사질 퇴적상으로 퇴적상이 변화하여, 민얼굴갯지렁이(S. bombyx)의 급격한 개체수 변동은 채집 지역의 퇴적상 변화와 관련이 높은 것으로 여겨진다.
긴자락송곳갯지렁이(L. longifolia)와 양손갯지렁이(M. japonica)는 니질퇴적물(고리) 또는 사질퇴적물(울진)로 되어진 발전소 취배수구 인근 지역인 거의 출현하지 않았고, 수심이 깊은 지역에서 우점하였다(Fig. 4). 긴자락송곳갯지렁이(L. longifolia)는 니질이 우세한 사·니질 혼합 퇴적상에서 출현하며 빈산소 수괴에서도 출현하는 잠재 유기물오염지시종으로 알려져 있다(Lim et al. 2006). 일본 세토내해에서는 사질 함량이 높고 수심이 낮은 해역에서 주로 출현하고 있다(Nishijima et al., 2015). 또한 양손갯지렁이(M. japonica)는 수심이 깊은 사질 퇴적상을 선호하는 종으로 알려져 있다(Kim et al., 2011). 저서동물의 종 조성은 퇴적물의 안정성과 매우 밀접하여, 퇴적물의 변동이 크면 종 조성 뿐만 아니라 개체수가 감소한다(Abe et al., 2015). 발전소 인근 취·배수구 지역에서는 높은 해수 유동증가로 인한 저서동물의 스트레스가 증가한다(Lardicci et al., 1999). 그러므로 본 연구에서 긴자락송곳갯지렁이(L. longifolia)와 양손갯지렁이(M. japonica)의 서식 환경이 양호한 사질 또는 니질 퇴적상에서 출현하지 않는 이유는 취·배수구의 해수 유동 증가에 의한 퇴적상의 변동이 커 개체수가 감소한 것으로 여겨진다. 오뚜기갯지렁이(S. scutata)는 고리의 대부분의 지역에서 많은 수로 우점 출현하였는데, 상당히 세립한 니질 퇴적상을 선호하는 오뚜기갯지렁이(S. scutata)의 특성 때문인 것으로 보인다(Shin et al., 1989). 버들갯지렁이과(Family Captelidae)의 Mediomastus californiensis도 고리의 몇 정점을 제외한 지역에서 많은 수로 출현하는 양상을 보였다. M. californiensis는 기회주의종으로 유기물오염이 진행되고 있는 해역에서 출현하는 유기물오염 지시종으로 알려져 있는데(Lim et al., 1991), 고리 주변 해역의 유기물 함량이 다른 해역에 비해 상대적으로 높아 M. californiensis가 우점하여 출현하는 것으로 여겨진다.
본 연구지역의 저서다모류 전체 군집 구조 분석 결과, 조사 지역은 비유사도 85.26%로 고리 지역인 (I)그룹과 울진, 후포 지역의 (II)그룹으로 크게 2개로 구분되었으며, 각각의 그룹은 각기 특징적인 환경상과 생물상을 보였다. 두 군집의 차이를 유발하는 가장 큰 요인은 퇴적물 내의 총 유기탄소량과 용존 산소량, 퇴적물의 입도와 퇴적상, 퇴적물의 분급도로 나타났다(Table 2, Fig. 6). 국내 연구의 경우 동해안 원자력 발전소 주변의 저서다모류 군집만을 대상으로 한 연구 결과가 없고, 대형저서동물의 군집 연구 결과도 많지 않아 직접 비교는 힘들지만 동해 후포주변 사질조하대에 서식하는 대형저서동물 군집은 수심과 퇴적물의 입도, 퇴적물 내의 총 유기탄소량에 영향을 받았다(Paik et al., 2007). 또한 동해 강릉 연안의 사질 퇴적물에 서식하는 대형 저서무척추동물의 분포는 수심에 큰 영향을 받으며(Choi et al., 2000), 동해 기장군 주변 해역에서 서식하는 저서다모류는 퇴적상에 따라 출현 종수 및 개체수의 차이가 나타났다(Kim et al., 2011). 특히, 원자력 발전소의 온배수가 대형저서동물의 군집에 영향을 미치는 것으로 알려져 있지만(Yu et al., 2011; 2013), 본 연구 결과 수온 변화가 저서다모류 군집에 미치는 영향은 없는 것으로 나타났다(Table 3). 그러므로 발전소 주변해역의 저서다모류의 군집구조에 영향을 주는 온배수 및 취·배수구의 인위적인 환경 교란은 발전소의 취·배수구 부근에 국한되며, 그 이외의 지역에서는 수심, 퇴적물 내의 유기 탄소 함유량(TOC)와 퇴적물의 평균 입도, 퇴적상 등 일반적인 환경 요인이 저서다모류의 종 조성과 군집구조에 더 중요하게 영향을 미치는 것으로 여겨진다.
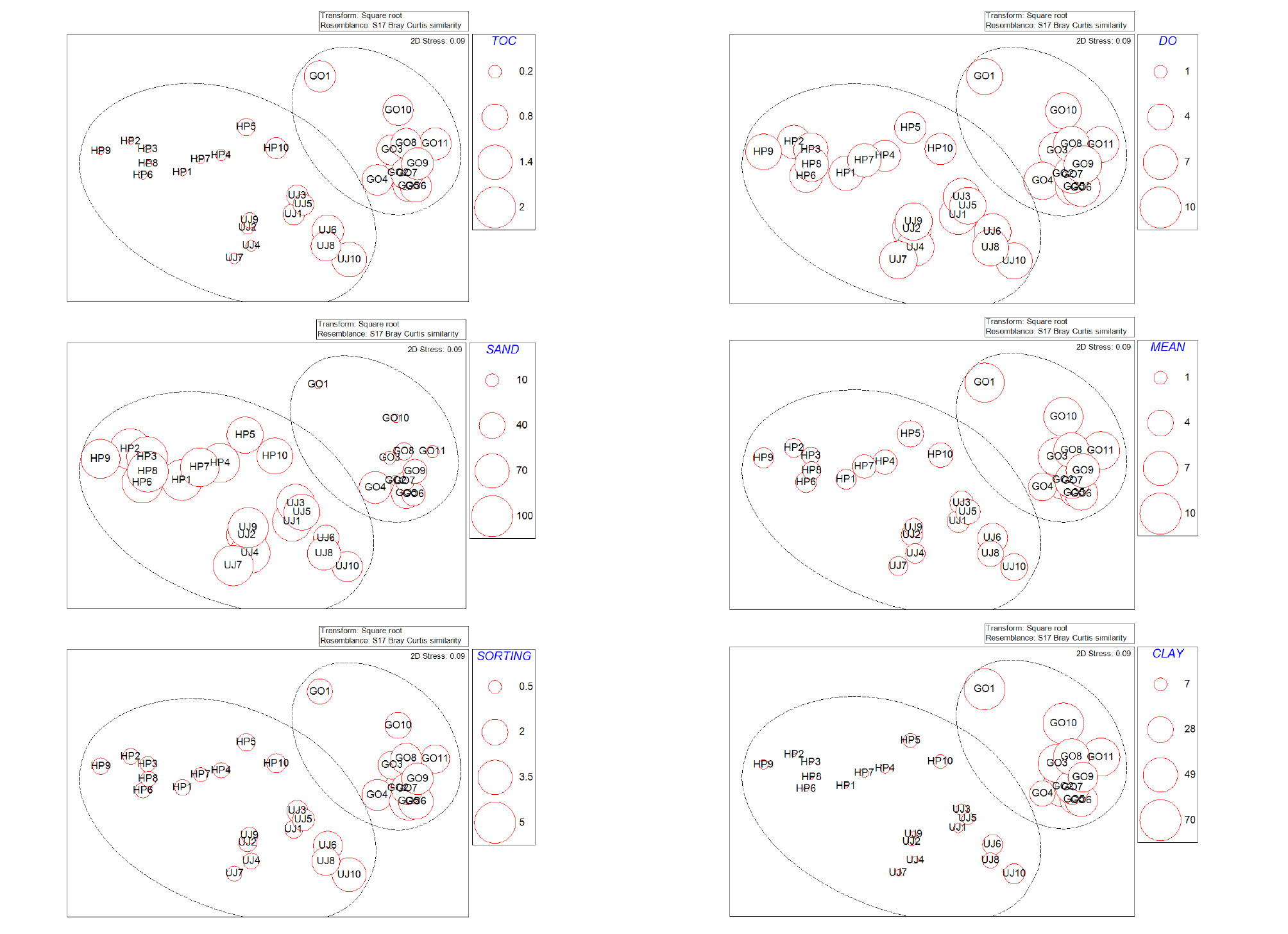
Fig. 6.
Multi-dimensional scaling (MDS) ordination showing the relations between benthic polychaete communities and four environmental factors such as total organic carbon (TOC), dissolved oxygen (DO), sand %, mean grain size (mean), sorting value of sediments (sorting), and clay % in the study area.
