

1. 서 론
대한해협은 쿠로시오의 지류인 대마난류와 황해 저층 냉수, 양자강 저염수, 다양한 연안수들이 유입되어 복잡한 수괴 구조가 형성되고, 이로 인한 생물 서식 환경에 영향을 줄 수 있다(Yang and Kim, 1990; Yang et al., 1998; Choo and Kim, 1998; 임 등, 2003). 다양한 경로를 통해 수괴와 함께 유입되는 질산염(NO3-)은 대한해협의 식물 플랑크톤 생산성 및 군집 구조의 변화를 일으킬 수 있다(Baek et al., 2010).
여름철 대한해협의 적조는 어업에 상당한 피해를 주고 있는데, 이에 대한 원인으로 동중국해에서 유입되는 고영양염의 수괴가 원인이 될 수 있다는 연구가 보고되었다(Lee et al., 2015). Lee et al.(2015)은 대한해협 적조 발생시 질산염이 제한인자임을 관측자료를 통해 밝혔다. 반면에 겨울철에 나타나는 질산염 고갈이 대한해협 서안에서 김 황백화 현상의 주요인이 될 수 있다고 선행 연구를 통해 제시되었다(NFRDI, 2014). 따라서 대한해협으로 유입되는 질산염의 기원을 파악하는 것은 해당 권역의 계절별 질산염 양극화 현상의 원인을 파악하고 대응하기 위한 필수적인 요소 중 하나라고 볼 수 있다.
대한해협으로 유입되는 해류는 제주 해협으로 유입되는 해류 및 제주도 동쪽과 일본 연안 사이로 유입되는 대마난류가 있고, 여름철 황해 내부에서 남진하는 황해저층냉수와 양자강 저염수, 대만 해협으로부터 기원하여 동중국해를 통과하는 대만난류수 등이 유입된다(Cho and Kim, 1994; Lee and Choi, 2009). 대한해협으로 유입된 수괴 및 해류는 대한해협 내부의 연안수와 만나 열염전선을 형성하며 대마도의 동쪽과 서쪽으로 나뉘어 동해 표층 및 중층으로 유입된다. 특히, 동해 중층으로 유입되는 대마난류 중층수는 영양염 농도가 높아 동해 내 일차 생산 및 생태 환경에 중요한 영향을 미친다(Rho et al., 2010; Rho et al., 2012).
선행 연구는 대마난류 중층수에 존재하는 영양염의 기원을 동중국해 및 대한해협에서 발생하는 재광물화 작용과 쿠로시오 해류로부터 기원할 수 있음을 제시하였다(Isobe, 1999; Kim et al., 2005; Guo et al., 2006; Cho et al., 2009; Morimoto et al., 2012; Kodama et al., 2015; Takikawa et al., 2016). Kim et al.(2013b)은 대한해협, 황해 및 동중국해에서 지속적으로 용존무기질소가 증가하고 있고, 동중국해로 유입되는 해류의 영양염 변화가 이에 대한 주 요인이라고 제안하였다. 또한, Chung et al.(2000)은 1997년부터 1999년까지 3년간 관측 조사를 통해 대한해협을 유출입하는 해류의 영양염 수송량을 비교 분석하였다. 선행 연구를 통해 대한해협의 질산염 변화는 해류 및 생지화학적인 과정을 통해 다양한 경로를 통해 발생함을 알 수 있었지만, 이에 대한 정량적인 평가와 비교 분석은 이루어지지 않았다.
본 연구에서는 3차원 물리-생지화학 결합 모델을 통해 대한해협 내 계절별 해류 및 생지화학과정을 통한 질산염 유출입량을 산출하고, 각 요인 별 질산염 유출입을 정량적으로 평가하고자 한다.
2. 방 법
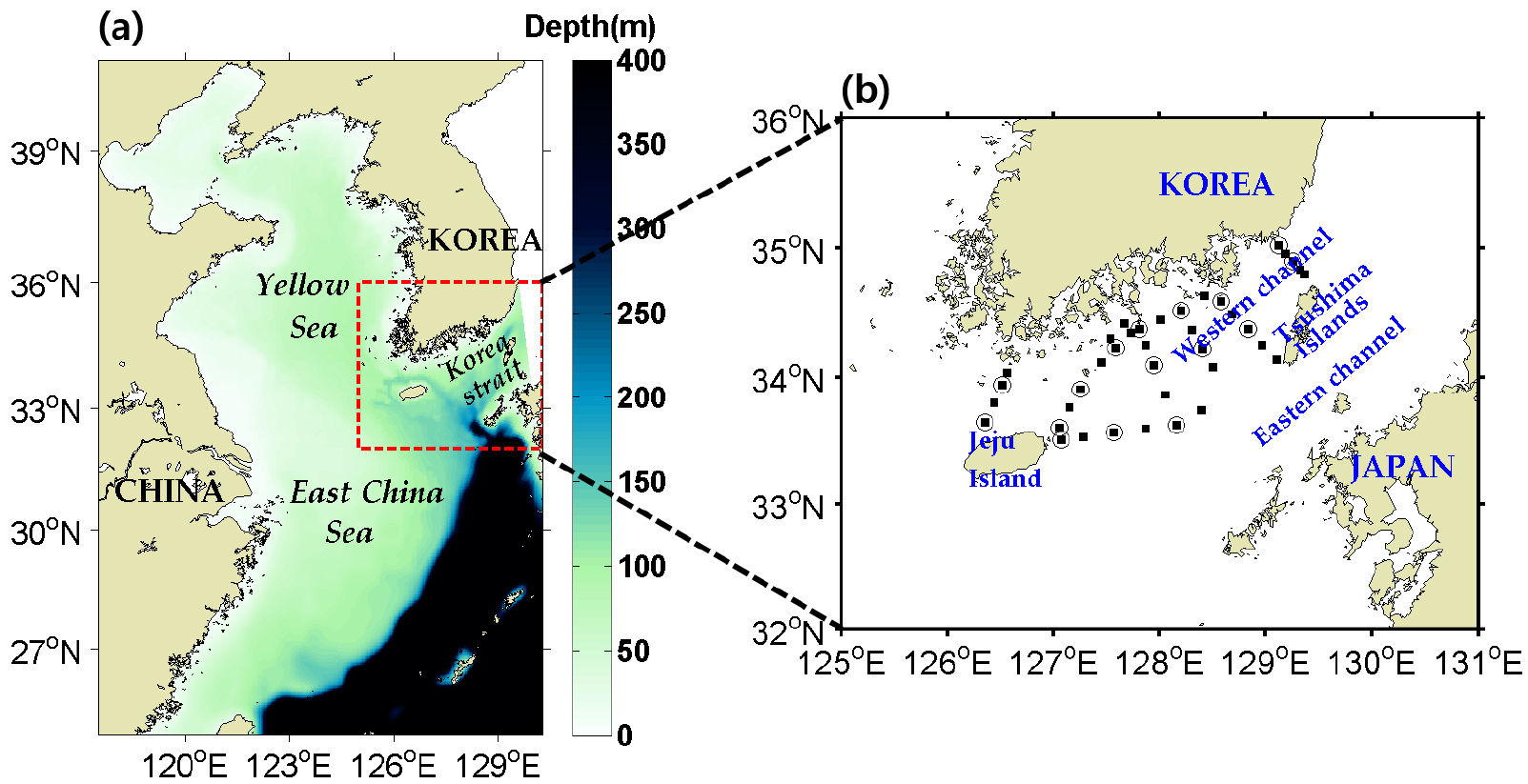
대한해협의 수온, 염분 및 질산염의 계절별 수평 및 수직 구조를 파악하기 위해 국립수산과학원의 2011년부터 2019년까지 204–207, 400 정선 자료를 이용하였다(Fig. 1b) 정선 자료는 2, 4, 6, 8, 10, 12월로 제공하고 있으며, 수심은 0, 10, 20, 30, 50, 75, 100, 125 m 자료를 제공한다. 본 연구에서는 표층과 저층의 대표 수심인 0, 50 m의 자료를 이용하였다. 자료 중 1 표준 편차 범위를 넘어서는 자료들은 분석의 정확도를 향상시키기 위해 제외한 후 분석에 실시하였다. 대한해협의 식물 플랑크톤 계절 변화를 분석하기 위해서 National Aeronautics and Space Administration에서 제공하는 Moderate Resolution Imaging Spectroradiometer (MODIS) 인공위성 기반 클로로필-a의 월 평균 자료를 이용하였다.
대한해협을 통과하는 해류와 생물학적인 요인에 의한 질산염 유출입을 정량적으로 평가하기 위해 물리-생지화학 결합 모델을 수행하였다. 모델은 Regional Ocean Modeling System (ROMS)모델을 이용하였다. ROMS는 수평적으로는 곡선 좌표를 수직적으로는 수심 변화를 반영할 수 있는 s-좌표계를 이용하며, 정수압, 부시네스크 근사를 통해 원시 방정식을 해석하는 모델이다(Shchepetkin and McWilliams, 2005). 본 연구를 수행하기 위해 수평 해상도는 1/30도 간격, 수직 해상도는 40개 층을 갖는 지역 모델을 구성하였고 모델 영역은 24.8°N–41°N, 117.5°E–130.3°E로 황해, 대한해협과 동중국해를 포함하는 범위로 구성되었다(Fig. 1a). 수심 자료는 Kim et al.(2013a)에서 사용한 모델의 수심 자료를 활용하였다.
물리 모델의 개방 경계 자료는 자료 동화를 수행한 북서태평양 모델의 결과 중 수온, 염분, 해수면, 유속 자료를 월 별로 입력하였고(Seo et al., 2014), 대기 강제력은 European Centre for Medium-Range Weather Forecasts 에서 제공하는 ERA5 재분석 자료 중 6시간 간격의 바람, 대기 온도, 해면 기압, 상대 습도 자료를 입력하였으며, 1일 간격의 단파 복사, 강수 자료를 모델에 입력하였다. 해면 열속은 입력된 대기 강제력을 bulk formula에 대입하여 계산하였다(Fairall et al., 1996). 조석은 TPXO6 모델로부터 획득한 10개의 분조를 개방 경계에서 입력하였다(Egbert and Erofeeva, 2002). 강물은 중국 및 한반도 주변의 12곳에서 입력되었고, 양자강 방류량은 다퉁 지점에서 관측된 자료를 이용하였고, 한반도 주변의 강물 자료는 국립환경과학원에서 제공하는 자료를 이용하여 월 평균 값을 산출하여 입력하였으며, 나머지 강들의 방류량은 선행 연구 자료를 이용하였다(Vörösmarty et al., 1996; Wang et al., 2008). 모델의 초기 입력 자료로는National Ocean Data Center 의 World Ocean Atlas 2013 (WOA2013)자료를 이용하였다(Locarnini et al., 2013; Zweng et al., 2013).
생지화학 모델은 질소 순환 및 인 순환을 재현함으로써 질산염속을 적합하게 재현할 수 있는 기법을 Fennel et al.(2006) 및 Gan et al.(2014)에서 개발한 모듈을 차용하였다. 본 모듈은 식물 플랑크톤, 클로로필-a, 동물 플랑크톤, 질산염, 암모늄(NH4+), 인산염(PO43-), 질소 및 인에 대한 큰 쇄설물과 작은 쇄설물들을 예측 변수로 포함하고 있다. 생지화학 모델을 수행하는데 이용되는 모수들은 Table 1에 제시되어 있으며 선행 연구에서 제안한 초기 설정 값과 해당 모델 영역을 수행하는데 이용한 설정 값을 각각 표기하였다(Tak et al., 2022).
Table 1.
Biogeochemical model parameters used in this study and default values from Fennel et al.(2006)
Fennel et al.(2006)에서 제안한 식물 플랑크톤 및 쇄설물의 침강 속도는 관측 자료 및 Stokes 수식, 모델 분석을 통해 일반적인 값으로 제안되어 있지만, 상수로 입력하게 되어있어 한계가 있다. 따라서 본 연구에서는 모델의 재현성을 높이기 위해 배경 수온의 변화에 따른 밀도 변화를 반영하여 침강 속도를 다르게 하여 모델을 수행하였다. Bach et al.(2012)는 수온이 10°C 이고 염분이 35 인 환경에서 수온이 25°C까지 오르면 입자성 유기물질의 침강 속도가 2배로 증가한다고 실험을 통해 밝혔다. 이를 토대로 해수 수온이 10°C보다 낮을 때 침강 속도는 균일하게 설정하고 10-25°C 구간에서는 선형적으로 침강 속도가 증가하며, 수온이 25°C 보다 따뜻할 경우에는 최대 침강 속도로 입자성 유기물질이 침강하게끔 설정하였다. 대한해협은 수온뿐만 아니라 염분도 계절에 따라 변한다. 하지만, 염분에 대한 침강 속도 변화를 살펴본 선행 연구가 부족하여, 대신 침강 속도가 수온에 따라 3배까지 증가할 수 있게 아래의 수식과 같이 수온에 따른 침강 속도 변화를 모델에 반영하였다.
여기서 는 수온을 나타내며, 와 는 수온에 따라 변하는 입자의 침강 속도, 초기 입력 침강 속도를 각각 나타낸다. 침강 속도가 수온에 따라 변하는 모듈을 통해 성층이 강한 여름철에는 입자성 유기물질이 표층에서는 좀 더 빨리 침강하고 수온이 낮은 저층에서 침강 속도가 느려져 수온약층 주위에 입자성 유기물질의 수렴을 실제와 가깝게 재현할 수 있었으며, 이를 통해 모델의 계절별 영양염 농도 분포를 이전 보다 개선 할 수 있었다.
생지화학 모델 변수 중 클로로필-a의 초기 입력 및 개방 경계 자료는 MODIS 인공위성 자료를 이용하여 경험식을 통해 산출하였다(Morel and Berthon, 1989; Uitz et al., 2006). 질산염과 인산염의 초기 입력 및 개방 경계 자료는 WOA2013 자료를 통해 획득하였으며, 암모늄은 일반적으로 농도가 매우 낮기 때문에 1 로 설정하여 모델을 수행하였다. 식물 플랑크톤과 동물 플랑크톤, 쇄설물의 경우 경험적으로 얻은 비율을 통해 산출하였다(Gruber et al., 2006). 양자강의 질산염, 암모늄, 인산염 농도는 2000년대 평균 값을 모델에 적용하였으며, 황하강 영양염 농도는 2002–2004년 평균 자료를 입력하였으며, 압록강 영양염 농도는 1992년, 1994년, 1996년 관측 자료의 평균 값을 입력하였다(Dai et al., 2011; Wang et al., 2011; Gong et al., 2015; Zhou et al., 2017). 한반도 연안에서 유입되는 한강, 금강, 영산강, 섬진강, 낙동강의 질산염, 암모늄, 인산염 농도는 국립환경과학원에서 제공하는 자료를 월 평균하여 입력하였다. 대기로부터 공급되는 질산염, 암모늄, 인산염 플럭스는 관측 자료를 토대로 0.027 , 0.080 , and 0.001 로 시공간적으로 균일하게 입력하였다(Zhang, 1994; Li et al., 2015).
물리-생지화학 결합 모델은 2006년의 개방 경계 및 대기 강제력으로 7년 동안 spin-up 기간을 거쳤으며, 이후 2019년까지 모델을 적분하였다. 적분 기간 중 2010년대 기간의 계절 변화를 분석하기 위해 2011년부터 2019년까지의 모델 결과를 분석에 이용하였다.
질산염 플러스를 산정하기 위해 아래와 같은 수식을 통해 물리적 요인과 생지화학적 요인을 구분지었다.
여기서 는 질산염 농도, 암모늄 농도, 식물 플랑크톤 농도, 유속 벡터, 질산염 농도의 확산 계수, 식물 플랑크톤의 질산염 섭취율(uptake rate), 질산화(nitrification) 계수를 각각 나타낸다. 우측 항에서 첫번째와 두번째 항은 이류 항과 확산 항으로 물리적인 요인으로 인한 질산염 변화를 나타내며 세번째와 네번째 항은 식물플랑크톤의 섭취와 질산화 작용에 대한 항으로 생지화학적 요인으로 인한 질산염 변화를 나타낸다. 여기서 확산에 의한 질산염 변화는 이류에 의한 변화보다 매우 적기 때문에 본 연구에서는 물리적인 작용에 대한 변화는 이류에 의한 변화만 취급하였다. 생지화학 모델에서 질산염 섭취율은 아래와 같은 수식으로 정의한다.
여기서 는 각각 식물 플랑크톤의 최대 성장률, 광합성-빛 관계식, 질산염 섭취에 대한 반포화 상수, 수온이 0°C 일 때 식물 플랑크톤의 성장률, 수온, 광합성이 반응하는 복사 에너지, 광합성 초기 기울기를 나타낸다. 는 수심(z)에 따라 기하급수적으로 감소하며 그 수식은 아래와 같다.
여기서 는 단파 복사 플럭스, 광합성 유효 복사 비율, 해수에 의한 빛 감쇄 계수, 클로로필-a농도로 인한 빛 감쇄 계수, 수심 𝜁 m 에서의 클로로필 농도를 각각 나타낸다. 는 아래와 같은 수식으로 산출된다.
여기서 , and 는 각각 최대 질산화율, 질산화 작용에 대한 빛 저해의 반포화 상수, 빛 저해 한계 수준을 나타낸다.
식 (2) 뿐만 아니라 생지화학적인 과정들과 생지화학적인 요소들에 대한 이류 및 확산은 모델의 모든 격자에서 산출된다. 1차 생산에 필요한 복사 에너지는 식 (6)에 의해 산출되며 각 격자의 수심별 클로로필-a 농도 및 해수의 빛 감쇄 계수를 반영하여 산출된다. 산출된 복사 에너지와 해당 격자의 수온 및 질산염, 암모늄, 인산염의 농도를 반영하여 1차 생산이 발생하며, 생산된 식물 플랑크톤은 식 (1)에 따른 침강 속도로 아래쪽 격자로 침강한다. 침강하면서 격자별 동물 플랑크톤의 농도에 따라 식물 플랑크톤은 섭식 되며, 일정 비율로 사망하여 유기 쇄설물로 변환된다. 유기 쇄설물은 동물 및 식물 플랑크톤의 사망으로 인해 생성되며, 일정 속도로 침강하면서 모델에서 설정된 재광물화율에 따라 무기 영얌염으로 변환된다. 재광물화 과정을 통해 유기물질은 인산염 및 암모늄으로 변환되며, 바닥층에 도달하는 일부 유기물질은 바닥층에서 즉시 재광물화 과정을 통해 인산염 및 암모늄으로 변환된다. 암모늄은 식 (7)과 같이 각 격자별 복사 에너지에 따라 질산화 과정을 통해 질산염으로 변환된다. 위에서 서술한 일련의 과정들은 수치 모델이 수행되는 60초 간격마다 격자별로 계산되며 본 연구에서는 산출된 값을 월 평균하여 분석을 실시하였다.
3. 결과 및 토의
물리-생지화학 모델 검증을 위해 국립수산과학원에서 제공하는 2월과 8월의 수온, 염분, 질산염의 표층 및 수심 50 m의 정선관측자료와 모델 결과를 비교하였다(Figs. 2, 3). 모델이 대한해협 환경의 계절에 따른 공간 분포를 적절히 모의하는지 확인하기 위해 모델 결과와 관측 자료 모두 2011년부터 2019년까지 월별 평균 자료로 재구성하여 비교하였다. 2월의 수온, 염분, 질산염 분포는 표층과 50 m 수심에서 유사하게 나타났는데, 이는 바람과 냉각에 의한 강한 수직 혼합 때문으로 보인다.
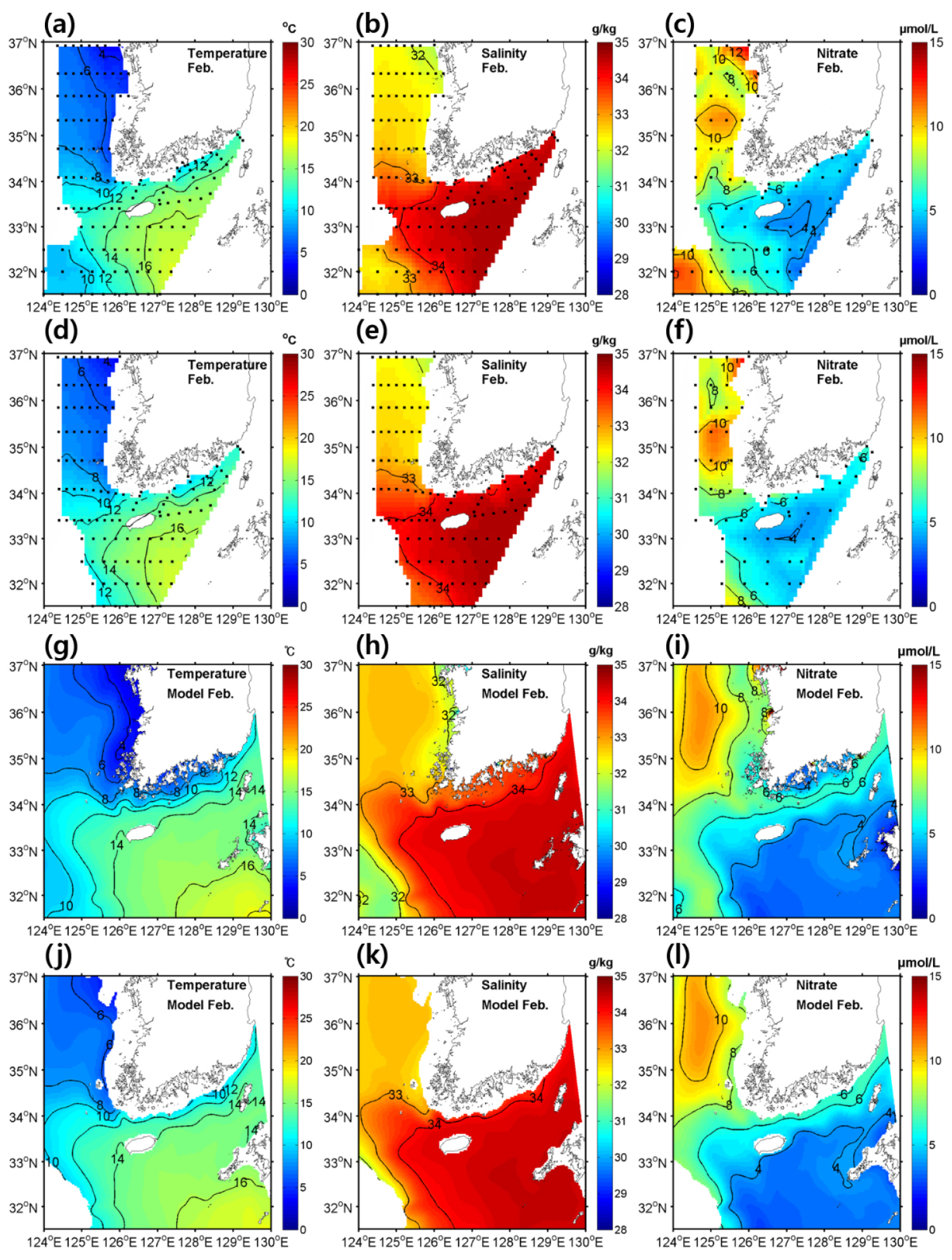
Fig. 2.
(a, d, g, j) Horizontal distributions of temperature (units are °C), (b, e, h, k) salinity (units are ), and (c, f, i, l) NO3- concentrations (units are ) from (a–f) observations and (g–l) model outputs at (a–c, g–i) the surface and (d–f, j–l) a depth of 50 m in February averaged for the period 2011–2019.
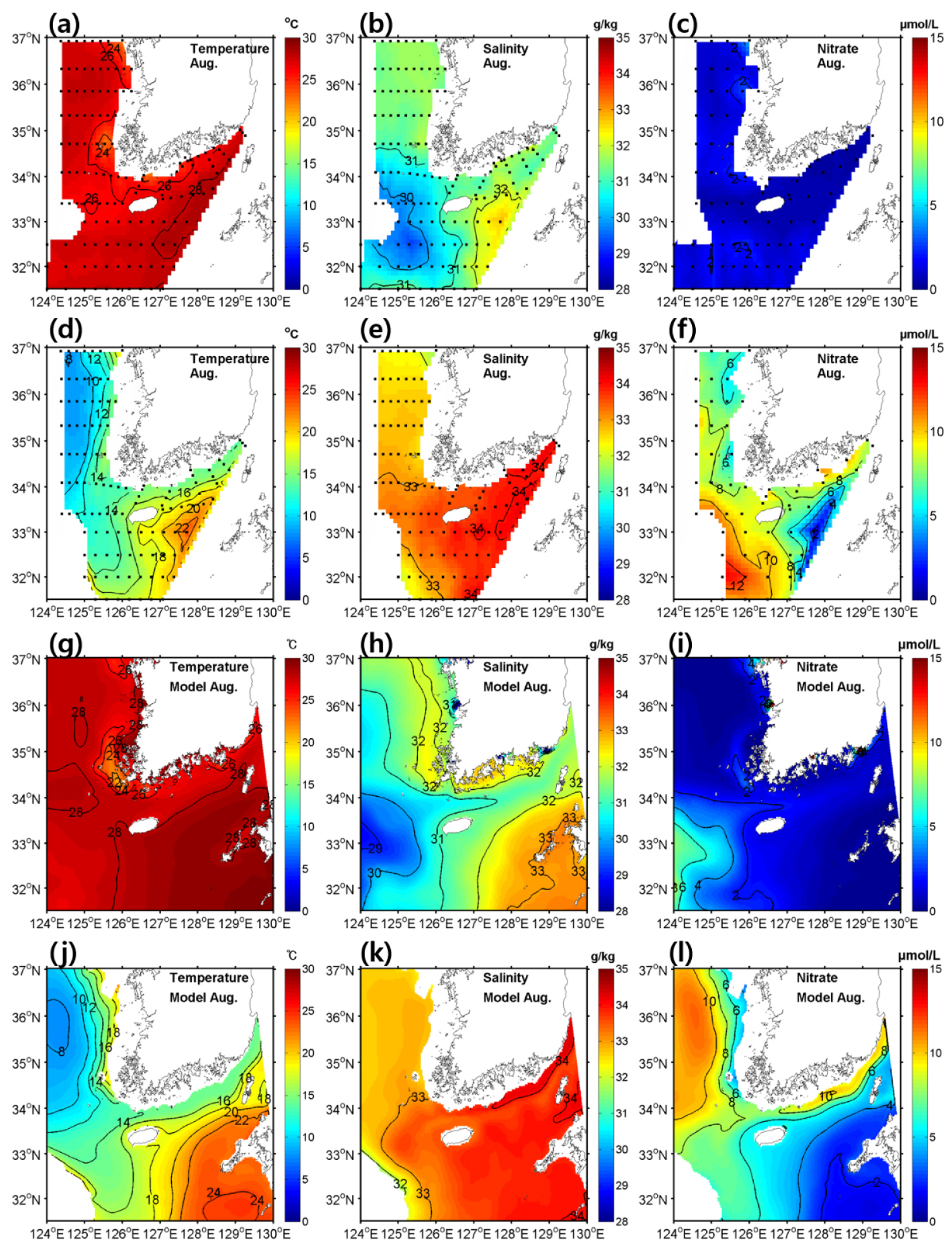
각 분포를 살펴보면 제주 서부에서 황해 중앙부로 고온, 고염이며 질산염 농도가 낮은 수괴가 황해난류로 인해 황해로 유입되는 것을 볼 수 있는데, 이러한 특성을 모델에서 적합하게 재현하고 있음을 알 수 있다(Tak et al., 2016). 8월에는 대부분의 표층 영역에서 수온이 26°C 보다 높게 나타났고 한반도 서쪽 및 남쪽 연안에서는 조석으로 인한 혼합으로 인해 상대적으로 수온이 낮아지는 것을 알 수 있었다(Sun and Cho, 2010). 수심 50 m에서는 황해 중앙부에 황해저층냉수라 불리는 냉수괴가 나타났다. 표층 염분 분포에서는 양자강 저염수로 인한 저염수괴가 제주도 서쪽 해역에서 나타났다. 질산염은 표층에서는 일차 생산으로 인해 전 영역에서 농도가 매우 낮았으며 수심 50 m에서는 대한해협과 제주도 서쪽에서 농도가 높았고, 제주도 동쪽에서는 상대적으로 농도가 낮게 나타났다. 8월의 모델 결과 또한 역시 2월과 마찬가지로 관측 자료의 수온, 염분, 질산염 분포와 유사하게 나타남을 알 수 있었다.
결합 모델이 생지화학적 요소들의 계절 변화를 적합하게 재현되었는지 판단하기 위해 질산염, 클로로필-a의 월별 시계열 분포를 관측 자료와 비교 분석하였다(Figs. 4a-c). 표층 질산염 농도는 봄철 대번성으로 인해 3월부터 6월까지 급격하게 감소하였고, 이후 가을, 겨울이 되면서 수직 혼합이 강해져 저층에 존재하는 질산염이 공급되면서 질산염 농도가 증가하였다. 수심 50 m에서 질산염 농도의 계절 변화는 표층에 비해 약하게 나타났으며 증감 형태 또한 다른 양상을 보였다. 봄철 대번성으로 인해 2월부터 4월까지 감소하는 경향이 나타났지만, 여름철에는 표층과 다르게 농도가 증가하는 형태를 보였다. 이후 가을에서 겨울로 접어들 때 수직 혼합이 강해지고 저층의 질산염이 표층으로 공급되면서 수심 50 m에서는 질산염 농도가 감소하였다.
클로로필-a는 질산염 농도가 감소하는 4월에 농도가 급격히 증가하는 형태를 보였고, 이후 농도가 줄어들다가 가을이 되면서 표층의 질산염 농도가 높아지기 시작할 때 가을 번성이 나타났다. 시계열 분석을 통해 계절 및 수심에 따른 질산염 농도 변화를 모델이 적합하게 재현하는 것을 알 수 있었다. 클로로필-a 농도는 겨울철을 제외하고는 모델이 다소 높게 재현하는 경향을 보였지만, 계절적인 특징은 잘 모의하는 것을 알 수 있었다.
물리-생지화학 모델 성능의 정량적인 평가를 위해 테일러 다이어그램을 통해 모델에서 재현된 수온, 염분, 질산염과 국립수산과학원의 관측 자료에 대해서 상관관계, 표준편차, 평균 제곱근 편차 분석을 표층, 수심 20 m, 수심50 m 에서 각각 실시하였고, 모델의 클로로필-a 자료는 MODIS 인공위성 자료와 비교 분석을 실시하였다(Fig. 4d). 클로로필-a와 수심 50 m의 질산염 농도의 상관관계가 각각 0.39와 0.46으로 상대적으로 낮게 나타났지만, 95% 신뢰구간에서 유의미한 값을 보였기 때문에 모델이 대한해협에서 발생하는 생지화학적인 과정을 적합하게 모의하였음을 알 수 있었다.
대한해협 주변의 해류는 계절에 따라 변하는 것으로 알려져 있다(Chung et al., 2000, Takikawa and Yoon, 2005). 모델이 대한해협 주변 해류의 계절 변화 모의 성능을 평가하기 위하여 대한해협 수송량을 관측 결과와 비교하였다. Takikawa and Yoon(2005)은 한국과 일본 연안의 해수면 차이를 통해 대한해협 수송량을 산정하는 경험식을 제안하였고, 이를 통해 산정한 대한해협 수송량과 모델 결과에서 계산한 대한해협 수송량을 비교해보았다(Fig. 4e). 대한해협 수송량은 겨울철에 2 Sv 정도로 낮게 나타나다가 늦여름, 가을철에 3 Sv 이상으로 최대치를 나타낸다. 모델 결과는 겨울철에 나타나는 최솟값은 해수면 차이를 기반한 경험식과 유사하게 나타났지만, 가을철에 나타나는 최댓값은 0.5 Sv 정도 낮게 모의하였다. 그림에는 표시되지 않았지만, 2011년부터 2015년까지 지속적으로 감소하는 경향도 모델에서 경험식과 유사하게 모의하였으며 상관계수 또한 0.84로 매우 높게 나타난 것으로 보아 모델이 실제 흐름을 적합하게 모의하고 있는 것을 알 수 있었다.
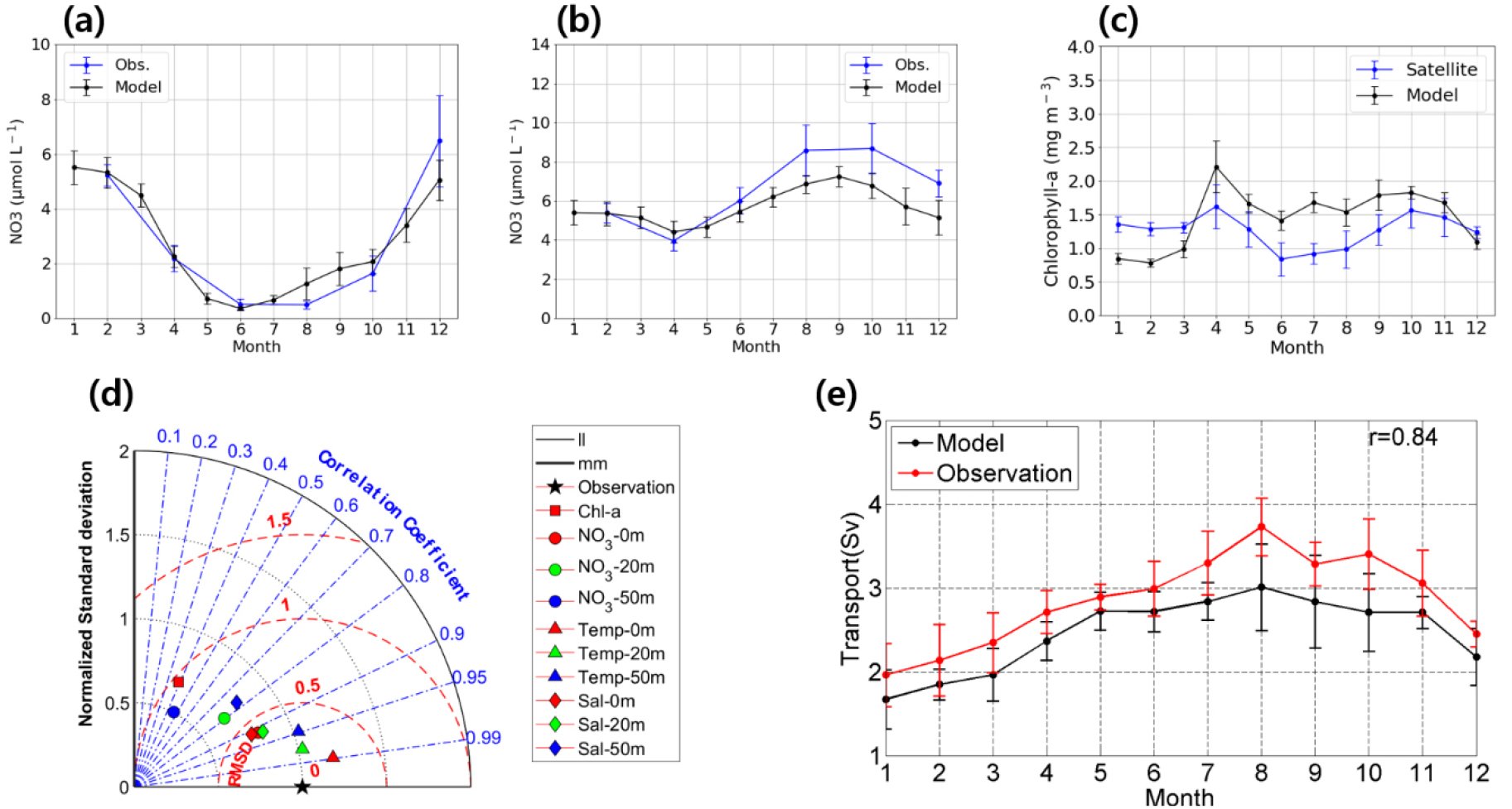
Fig. 4.
Time-series of climatological monthly mean of NO3- concentrations (units are ) at (a) the surface layer and (b) a depth of 50 m and (c) chlorophyll-a concentrations (units are ) from observations, satellite data (blue line) and model outputs (black line). Chlorophyll-a concentrations are averaged over the south coastal sea of Korea (126°E–129°E, 33.5°N–35°N) from 2011 to 2019. Vertical bars denote the standard deviation of interannual variation. (d) Normalized Taylor diagram comparing simulated chlorophyll-a concentration (squares) averaged over the region same as (c) and temperature (triangles), salinity (diamonds), NO3- concentration (circles) at the surface (red), 20 m depth (green), 50 m depth (blue) averaged at stations indicated in Fig. 2b with observation, satellite data (the reference field indicated by a black star on the x-axis). The radial coordinate presents the magnitude of normalized standard deviation from observation. The angular coordinate indicates the correlation coefficient (denoted by blue dash-dotted lines). The concentric semi-circles (red) indicate Root Mean Square Difference (RMSD). (e) Time-series of climatological monthly mean volume transport (units are ) through the Korea Strait averaged from 2011 to 2015 using an empirical equation (red line) and model outputs (black line). Vertical bars denote one standard deviation.
검증이 이루어진 모델 결과를 토대로 격자별 유속과 이로 이한 질산염 수송의 계절 변화를 표층과 저층으로 나누어 살펴보았다(Fig. 5). 봄, 여름, 가을, 겨울철 평균 질산염 수송은 3–5월, 6–8월, 9–11월, 12, 1, 2월의 평균치로 정의한다. 표층과 저층은 에크만 수심을 고려하여 표층은 해표면부터 수심 30 m 까지를 평균한 값이며 저층은 수심 30 m 부터 바닥까지 평균한 값으로 나타내었다. 유속에 의한 표층 질산염 수송은 주로 제주해협으로부터 유입되는 양이 우세하게 나타났으며 겨울, 봄철에는 한국 서부연안에서부터 연안을 따라 반시계 방향으로 흐르는 연안류에 의한 수송이 강하게 나타나는 것을 알 수 있었다. 유속 분포와는 다르게 여름, 가을에는 제주도와 큐슈 사이로 유입되는 질산염 수송은 거의 존재하지 않았는데, 이는 해당 권역의 여름철 표층 질산염 농도가 매우 낮기 때문인 것으로 보인다(Fig. 3i). 반대로 저층의 질산염 수송은 제주도와 큐슈 사이로 유입되는 것이 매우 크게 나타났다. 이는 대마난류 중층수에 해당하는, 수온은 12-15°C 이며 염분은 34.3 이상인, 고온 고염의 질산염 농도가 높은 수괴가 동중국해를 통과하여 유입되기 때문인 것으로 보인다(Kim et al., 2007; Rho et al., 2010). 유속 분포는 표층이 중층보다 강하게 동쪽으로 흘러갔지만, 질산염 농도가 저층이 더 높기 때문에 질산염 변화는 저층에서 더 높게 나타났다.
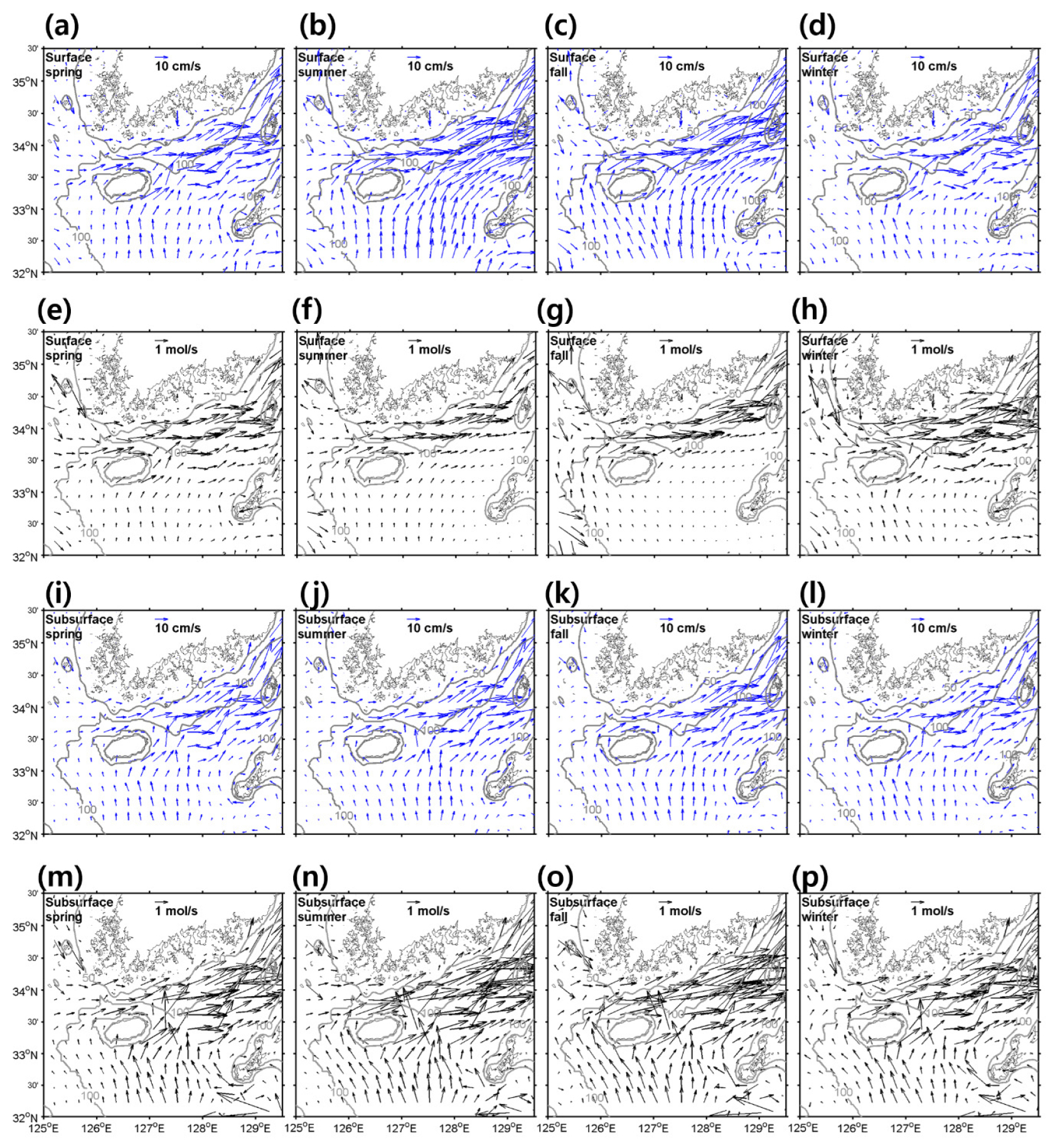
Fig. 5.
Horizontal distributions of velocities (blue arrows) at (a–d) the surface, (i–l) subsurface layers, NO3- transports by currents (black arrows) at (e–h) the surface and (m–p) subsurface layers with (a, e, I, m) spring, (b, f, j, n) summer, (c, g, k, o) fall, and (d, h, l, p) winter. The surface and subsurface layers are averaged from the surface to a depth of 30 m and from 30 m to the bottom. Grey contours indicate depths of 50, 100 m. Units are .
생지화학적 과정을 통한 질산염 유출입의 계절에 따른 공간적인 변화를 조사하기 위해 식물 플랑크톤의 질산염 소비와 질산화작용에 의한 공급 분포를 나타내었다(Fig. 6). 질산염 섭취는 봄철 대번성으로 인해 봄에 가장 높게 나타났고 가을에 가장 낮게 나타났다. 여름철에는 조석으로 인한 혼합으로 표층에 질산염이 공급되면서 연안쪽에서 섭취가 강하게 발생하였다. 질산화 작용은 제주도 서남쪽과 황해해역에서 0.2–0.4 정도 나타났지만 대한해협에서는 그 이하로 매우 낮게 나타났다. 식물플랑크톤의 질산염 섭취로 인한 소비와 질산화 작용으로 인한 생산의 차이를 살펴보면 연안에서는 소비가 크게 우세하였으나 외해에서는 생산이 약간 우세하게 나타났다.
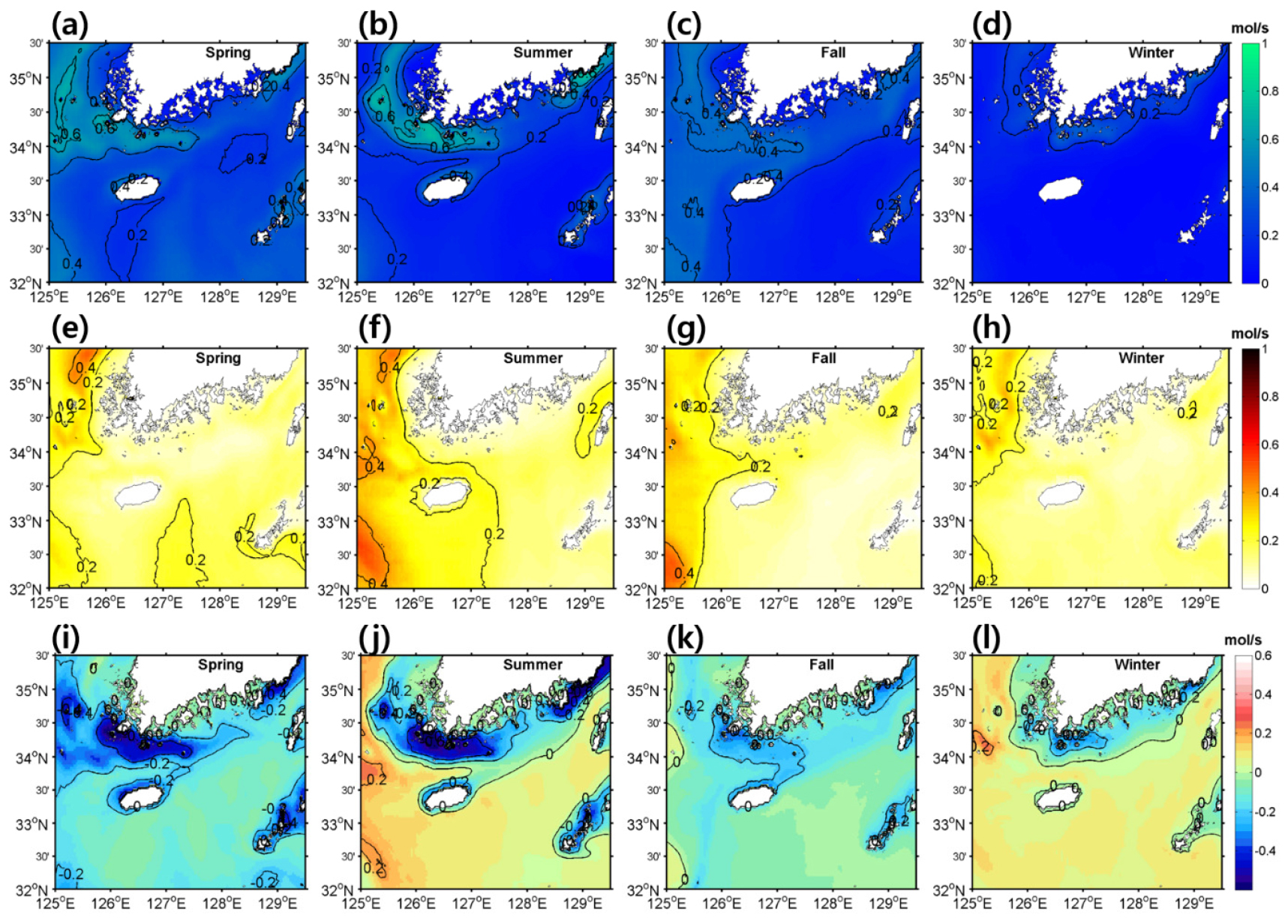
Fig. 6.
Horizontal distributions of vertically integrated (a–d) uptake of NO3-, (e–h) nitrification, and (i–l) difference between uptake and nitrification with (a, e, i) spring, (b, f, j) summer, (c, g, k) fall, and (d, h, l) winter. Positive and negative values in the difference indicate net supply and consumption by biogeochemical process. Units are .
정량적인 평가를 위해 해류와 대한해협에서 발생하는 생지화학적인 요인들에 의한 질산염 유출입을 월별 시계열 변화를 통해 살펴보았다(Fig. 7). 해류에 의한 유입을 살펴보면 제주도와 일본 큐슈 사이로 유입되는 대마난류에 의한 질산염 수송이 가장 많음을 알 수 있었으며 8월에 가장 높은 값을 보였다(Fig. 7b). 이 중 일부는 대한해협 동수도를 통해 동해로 유입되었는데, 동수도에서 나타난 질산염 수송의 최댓값은 3월에 나타났다. 대마난류에 의한 질산염 유입량 중 동수도로 유출되지 않은 나머지와 제주해협을 통한 질산염 유입량은 서수도로 유출되었고, 이들의 최댓값은 공통적으로 가을철에 나타났다. 제주해협으로 유입되는 질산염은 연 평균 2.5 로 나타났고, 대한해협을 통해 유출되는 질산염 수송량은 9.7 로 선행연구에서 산출한 10.7–13.2 과 유사한 수치를 보였다(Kim et al., 2013b; Chung et al., 2000). 제주도와 큐슈 사이로 대마난류에 의해 유입되는 질산염 수송량은 연 평균 7.6 로 이 또한 선행연구에서 제시한 수치인 8.0 와 유사한 수치를 나타내었다.
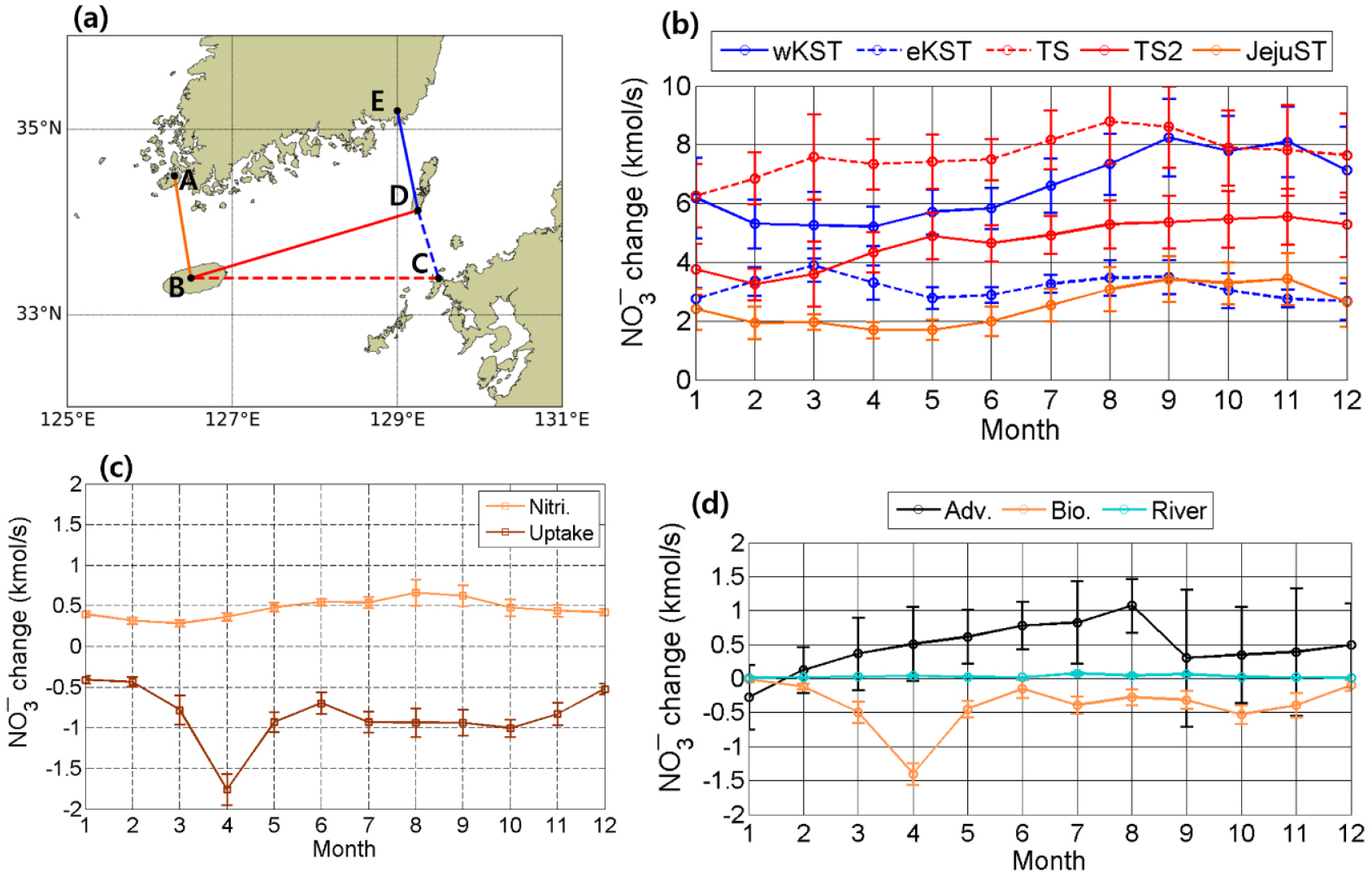
Fig. 7.
(a) A region to estimate NO3- change based on currents across sections indicated by colored lines and NO3- uptake and nitrification. (b) Monthly mean net advective NO3- change across sections of A–B (orange), B–C (red dashed), C–D (blue dashed), D–E (blue), B–D (red). Positive values are eastward and northward transports. (c) Monthly mean NO3- change by the uptake (brown) and nitrification (orange) in a box surrounded by A, B, C, D, E in (a). (d) Monthly mean net NO3- change by currents (black) and biogeochemical processes (orange) in the box used in (c). Cyan line indicates NO3- input by Sumjin river and Vertical bars indicate the standard deviation of interannual variation.
연 평균 질산염 수송량을 통해 각 해협으로 유출입되는 양을 평가해보면, 제주도와 큐슈 지역 사이를 통해 대마난류로 유입되는 수송량은 제주해협을 통한 유입량 보다 3배 많았으며, 대한해협 서수도를 통해 유출되는 질산염 수송량은 동수도를 통해 유출되는 것보다 2배 더 많았다.
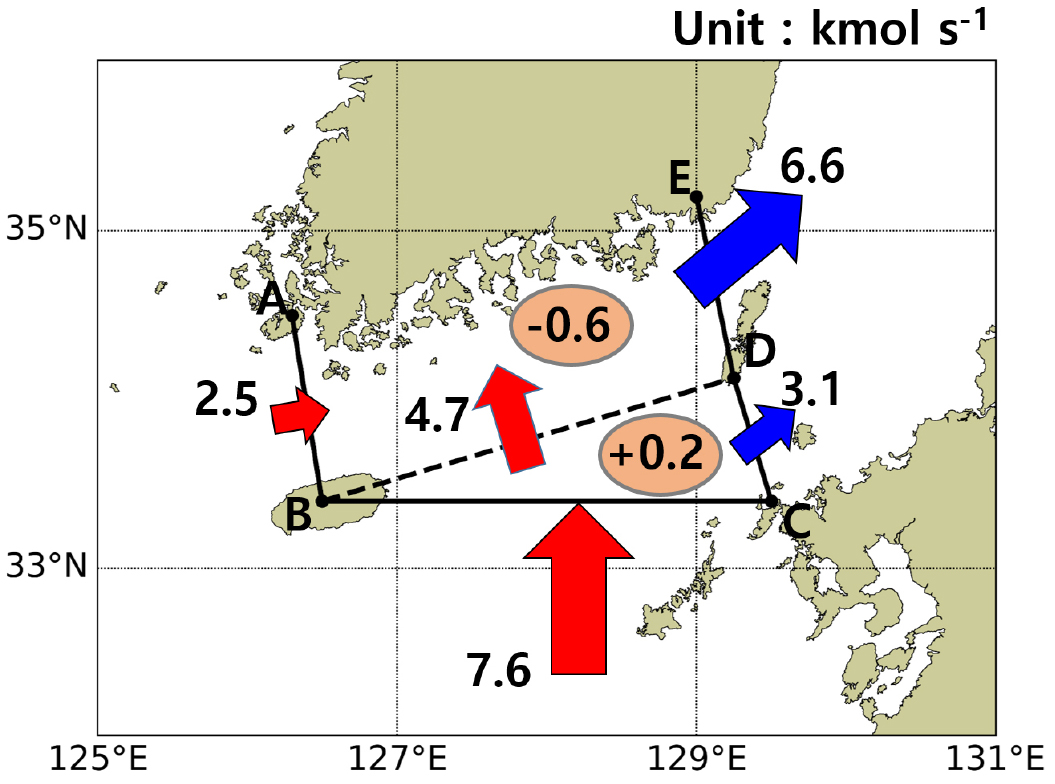
Fig. 7a에서 점 A, B, C, D, E로 둘러싸인 해역에서 봄철 및 가을철은 식물 플랑크톤의 번성으로 인해 질산염 소비가 많이 발생하였으며, 질산화 작용으로 인한 질산염 공급은 평균 0.5 정도로 연중 균일하게 나타났다. 대한해협에서 해류 및 생지화학적인 요인들로 인한 질산염의 순 변화량은 1월을 제외한 나머지 월에는 해류로 인해 항상 공급되는 것을 알 수 있었고, 공급된 질산염들은 겨울을 제외한 나머지 계절에 식물 플랑크톤으로 인해 소비되고 있음을 알 수 있었다. 강물에 의한 유입은 무시할 수 있을 만큼 적었다. 이를 통해 해류로 인해 대한해협으로 질산염이 공급되며, 해류에 의한 순 공급량은 식물 플랑크톤에 의해 소비됨을 알 수 있었다. 해류에 의한 유입량은 제주해협으로 유입되는 양보다는 대마난류의 중층부를 통해 유입되는 양이 3배 정도 더 많은 것을 알 수 있었으며, 이 중 1/3 은 대한해협을 경유하지 않은 채로 동수도를 통과하여 동해로 유입되었고, 2/3는 대한해협을 경유하여 서수도를 통과하여 동해로 유입됨을 알 수 있었다. 해류 및 생지화학적인 요인에 의한 질산염 유출입량의 모식도는 Fig. 8과 같다. Fig. 8에서 생지화학적 요인에 의한 질산염 변화의 경우, 대한해협 내 연안에 A-B-D-E 해역은 1차 생산에 의한 질산염 소비가 더 크게 발생하여 연 평균적으로 0.6 만큼 소비가 발생하였고, 상대적으로 외해에 해당하는 B-C-D 영역은 식물 플랑크톤에 의한 질산염 섭취가 적어 질산화 작용으로 인한 공급이 0.2 만큼 더 많았다.
4. 요약 및 제언
물리-생지화학 결합 모델을 이용하여 대한해협에서 해류 및 생지화학적 요인에 의한 질산염의 계절별 공간 변화를 계산하였다. 관측 및 인공위성 자료와 비교를 통해 결합 모델이 수온, 염분, 질산염, 클로로필-a의 계절 변동을 적합하게 재현하는 것을 알 수 있었다.
수치 모델 결과를 통해 해수유동에 의한 질산염 수송은 연중 같은 방향으로 유출입 되었다. 유입은 제주해협과 제주도와 일본 큐슈지역 사이로 유입되었으며, 대한해협을 통해 유출되었다. 표층에서는 봄, 겨울철에 한국 서부 연안에서부터 대한해협으로 흐르는 연안류를 통해 유입되는 양이 많았고, 저층에서 제주도와 큐슈 지역 사이로 유입되는 양이 많았다. 생지화학적인 요인에 의한 질산염 변화는 주로 연안에서 식물 플랑크톤에 의해 소비되는 모습이 연중 나타났다. 여름철에는 일반적으로 표층에서는 영양염 고갈으로 인해 식물 플랑크톤의 섭취율이 낮은데, 대한해협 연안부에서는 조석에 의한 혼합으로 인해 저층의 질산염이 공급되면서 소비가 지속적으로 나타났다. 질산화 작용에 의한 질산염 공급은 황해와 제주 서남부 해역에서 상대적으로 많았지만, 대한해협에서는 0.2 이하로 매우 낮게 나타났다. 해류에 의한 가장 큰 질산염 유입원은 제주도와 큐슈 지역 사이로 유입되는 대마난류에 의한 수송이었고, 제주해협을 통해 유입되는 양보다 대략 3배 정도 많은 양이 유입되었다.
대마난류를 통해 유입된 양 중 1/3은 대한해협 동수도를 통해 동해로 유입되었고, 나머지 2/3는 대한해협을 경유하여 대한해협 서수도를 통해 동해로 유입되었다. 각 요인에 대한 질산염 순 변화를 통하여 해류에 의한 질산염은 1월을 제외한 나머지 월에는 항상 대한해협에 질산염을 공급하였고 생지화학적인 요인으로 인해서는 겨울을 제외한 모든 계절에 소비됨을 알 수 있었다.
결합 모델과 결과 분석을 통해 대한해협으로 유출입하는 해류 및 생지화학 요인들로 인한 질산염 변화를 정량적으로 평가할 수 있었다. 하지만, 이밖에도 지하수 유입과 대기에 의한 질산염 유입에 대한 시공간적인 변화 또한 질산염 유출입에 중요한 영향을 줄 수 있다. 이 부분에 대한 추가적인 연구를 통해 모델의 정확도를 높인다면 추후 정확한 질산염 유출입에 대한 평가를 할 수 있을 것으로 판단된다. 본 연구에 이용된 경계 자료들이 주로 기후학적 평균 자료를 이용하였기 때문에 대한해협 질산염 변화에 대한 장기적인 경향을 분석하기에 어려움이 있었다. 추후 이런 부분도 고려하여 대한해협을 통과하는 해류들에 의한 질산염 수송이 장기적으로 어떤 영향을 미칠 지에 대해서도 연구해 볼 예정이다. 또한, 저차 생산 및 섭식이 고려된 생지화학 모델을 토대로 하여 고차 소비자의 섭식 및 호흡에 대한 부분을 반영한 대한해협의 생태계 변화 연구도 향후 중요한 연구 과제 중 하나이다.
