

1. 서 론
2. 자료 및 방법
2.1 O2/Ar을 이용한 혼합층 순군집생산 추정
2.2 18O 배양법을 이용한 유광대 총일차생산과 순군집생산 추정
3. 결 과
3.1 표층의 용존 기체 분포
3.2 유광대 총일차생산과 순군집생산
4. 토 의
4.1 연안과 울릉분지의 생산력 차이
4.2 연안과 울릉분지의 CO2 흡수력
4.3 연안의 ∆N2/Ar 변화와 질소 고정 가능성
5. 결 론
1. 서 론
해양 일차 생산자에 의해 광합성으로 고정된 유기 탄소가 심층으로 제거되는 과정을 생물 펌프(biological pump)라 한다(Volk and Hoffert, 1985). 이는 표층 이산화탄소 분압을 낮추어 대기 이산화탄소 분압을 조절하는 중요한 기작이다. 동해는 육상에서 영양염 공급을 많이 받는 대륙주변해로 생물 생산이 높고, 대기 이산화탄소의 제거원으로 알려져 있다(Kim et al., 2014). 동해의 주요 세 분지인 울릉분지, 일본분지, 야마토분지 중에서도 울릉분지는 가장 생산력이 높은 것으로 보고되었다. Yamada et al.(2005)은 위성 해색 자료를 활용하여 울릉분지의 순일차생산(총광합성과 독립영양생물 호흡의 차이; net primary production; NPP)이 222 g C m−2 y−1로 일본분지의 161 g C m−2 y−1과 야마토분지의 191 g C m−2 y−1보다 높은 것으로 추정하였다. Kwak et al.(2013)은 여름철 울릉분지의 순일차생산이 0.37 – 0.96 g C m−2 d−1 (n=5) 범위로 인접한 동중국해와 서태평양보다 높은 것으로 보고하였는데, 유광대 하부에서 공급된 질산염이 그 이유인 것으로 발표하였다.
총광합성과 군집 호흡의 차이로 정의되는 순군집생산 혹은 유광대 하부에서 공급된 질산염에 의한 신생산(new production; NP)은 생물 펌프를 평가하는 척도이다. Hahm et al.(2019)은 여름철 용승의 영향을 받은 해역의 순군집생산이 0.66 g C m−2 d−1로, 주변 해역에 비하여 두 배 이상 증가한 결과를 발표하였다. Kwak et al.(2013)은 여름철 울릉분지의 신생산이 0.15 – 1.3 g C m−2 d−1 (n=5) 범위로 0.01 – 0.03 g C m−2 d−1 (n=4) 범위의 동중국해 및 서태평양보다 높은 것으로 보고하였다. 높은 생물 생산은 용승, 에디, 해류 등과 같은 물리 요인과 함께 울릉분지 표층의 fCO2 분포를 조절하는 주요 요인이다. Kim et al.(2014)은 울릉분지의 월별 표층 fCO2 변화 요인을 수온 변화와 생물 활동으로 나누어 확인하였는데, 봄과 초여름(3-6월) 동안에는 생물 생산의 영향이 크게 작용하여 해수의 fCO2가 감소한다고 발표하였다.
연안은 전 지구 관점에서 대기 이산화탄소를 흡수하는 것으로 평가된다(Ducklow and McCallister, 2004). 그럼에도 불구하고 울릉분지에 비하여 동해 연안의 생물 펌프 및 탄소 흡수력에 대한 이해는 매우 제한적이다. Kwak et al.(2013)은 여름철 연안(35.5-37.5°N, <129.7°E)의 순일차생산은 0.45 – 2.41 g C m−2 d−1 (n=5) 범위로, 신생산은 0.42 – 5.95 g C m−2 d−1 (n=5) 범위로 보고하였는데, 공간 변동성이 큰 탓에 지속적인 현장 관측이 요구된다. 본 연구에서는 선상 연속 관측 시스템을 이용하여 표층의 ∆O2/Ar과 fCO2의 변화를 넓은 시공간 해상도로 관측하였다. 이들 용존 기체 포화도의 변화를 이용하여 그동안 많은 연구가 수행되지 않은 동해 연안의 생물 펌프 및 탄소 흡수력을 울릉분지와 비교하였다. 또한 울릉분지와 연안에서 배양 방법으로 일차 생산을 추정하여 두 공간의 생산력을 비교하였다. 마지막으로 연안에서 상당한 질소 고정 가능성을 시사하는 질소불포화 관측 결과를 함께 제시하였다.
2. 자료 및 방법
2.1 O2/Ar을 이용한 혼합층 순군집생산 추정
2.1.1 표층 용존 기체 관측
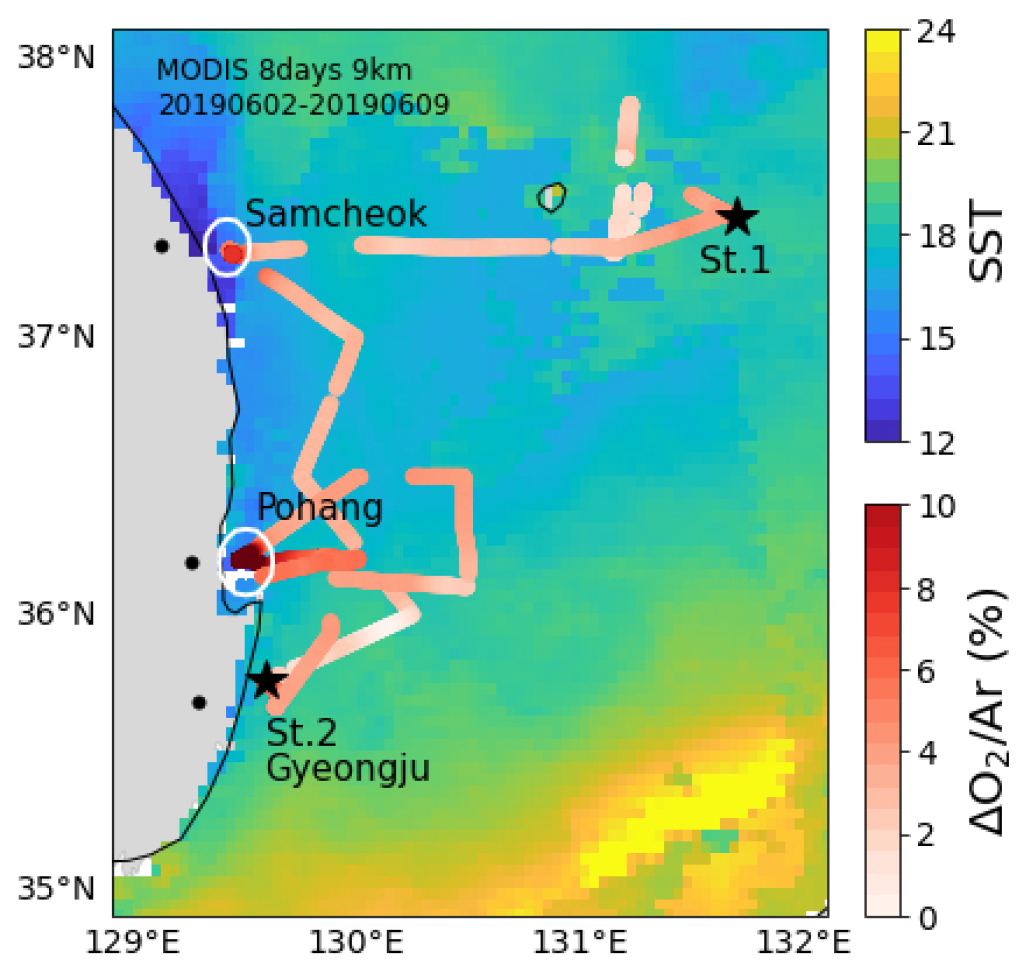
2019년 6월 5일에서 6월 10일까지 연구선 아라온 호에 승선하여 동해 남서부해역 표층의 용존 기체 분포 변화를 연속 관측하였다(Fig. 1). 약 7 m 수심에 위치한 연구선의 청정 해수 공급 장치를 이용하여 표층수를 실험실로 공급하였다. 표층수의 O2/Ar은 평형기 주입식 질량분석기(Equilibrator Inlet Mass Spectrometer, EIMS; Cassar et al., 2009; Hahm et al., 2014)를 이용하여 연속 관측하였다. 연구선 선수에서 채취된 깨끗한 대기를 표준 기체로 이용하였고, 약 3시간 간격으로 반복 측정하였다. EIMS로 측정한 표준 기체의 O2/Ar 변동성은 0.5%보다 작았다.
표층의 수온(SST)과 염분(SSS)은 Sea-Bird사의 thermosalinograph (SBE45)를 이용하여 관측하였다. 식물플랑크톤의 생물량을 대변해주는 지표인 표층의 엽록소 형광은 Turner Designs사의 10-AU fluorometer를 이용하여 관측하였다. 표층의 O2 농도는 Aanderaa사의 oxygen optode 4531을 이용하여 관측하였다. 제조사에서 제공하는 보정식에 따라 센서에서 얻어진 용존 O2 농도에 염분 보정을 하였고, Johnson et al.(2015)을 따라 득률 보정(gain correction)하여 최종 용존 O2 농도를 결정하였다. 득률 보정을 위해 optode로 매일 1 시간씩 실험실 내 공기의 O2 분압을 측정하였다. optode로 측정된 O2 분압과 실험실의 대기압과 온도로 계산한 O2 분압의 비로 득률을 결정하였다. 표층 해수의 pCO2는 General Oceanics사의 CO2 측정 시스템(Model 8050)을 이용하여 관측하였다. 측정된 pCO2로부터 Pierrot et al.(2009)에 따라 fCO2로 산출하였다.
2.1.2 혼합층 순군집생산 계산
순군집생산(net community production; NCP)은 총광합성과 총호흡의 차이로 정의된다. 일차 생산으로 고정된 탄소의 양에 비례하는 산소가 부산물로 만들어지기 때문에, 생물 군집에 의한 순산소변화를 정량함으로써 순군집생산을 추정할 수 있다. 그러나 해수 속의 O2 농도는 광합성과 호흡의 생물 현상 뿐만 아니라 수온 변화, 기체 용해, 기체 주입, 혼합 등의 물리 현상의 영향이 더해진 결과이다. 물리 현상에 의한 O2 농도 변화를 제거하기 위해 Ar을 함께 측정한다. Ar은 화학, 생물 과정에 반응하지 않는 비활성 기체로 용해도와 확산 계수가 O2와 매우 유사하여 물리 현상에 의한 O2 농도 변화를 나타내는 지표이다(Craig and Hayward, 1987). 생물학적인 산소 농도([O2]bio)는 다음 식으로 추정한다(Reuer et al., 2007).
여기서 [Ar]은 시료의 Ar 농도, [Ar]sat과 [O2]sat은 포화 상태의 Ar과 O2 농도이다. 생물 활동에 의한 산소 과포화(biological oxygen supersaturation)를 나타내는 ∆O2/Ar은 다음과 같이 정의된다.
여기서 ([O2]/[Ar])sample과 ([O2]/[Ar])sat은 각각 시료와 포화 상태의 O2와 Ar 농도 비이다. 연직 혼합과 수평 이류를 무시한다면, NCP (mmol O2 m−2 d−1) 는 [O2]bio가 기체 교환을 통해 대기로 빠져나가는 양과 같다. 이렇게 계산된 혼합층 순군집생산은 다음 절에서 기술하는 18O 배양법으로 추정한 유광대 순군집생산과 구별하기 위해 NCPin situ로 부르겠다.
여기서 [O2]sat은 대기와 평형 상태일 때 O2 농도(mmol kg−1)이고, ρ는 표층 해수의 밀도(kg m−3)이다. Ar은 대기와 평형 상태라고 가정하였다(즉, [Ar]/[Ar]sat=1). kw 는 일별 가중치를 둔 산소의 기체 교환 속도(m d−1)로 관측 시점으로부터 30일 이전까지의 풍속이 기체 교환에 미치는 영향을 고려하기 위하여 사용되었다(Reuer et al., 2007). 일별 가중치는 매일의 기체 교환 속도와 환기된 혼합층 비율로 결정되었다. 일별 기체 교환 속도 계산에는 일별 NCEP (National Centers for Environmental Protection) 재분석 자료에서 얻은 해수면 위 10 m 풍속 자료와 Wanninkhof(2014)의 풍속-기체 교환 관계식을 이용하였다. 혼합층 수심으로는 장기 평균 혼합층 수심 자료를 사용하였다(Levitus and Boyer, 1994). [O2]sat과 [Ar]sat은 각각 Garcia and Gordon(1992)과 Hamme and Emerson(2004)의 용해도 식으로 계산하였다.
2.1.3 이산화탄소 플럭스 계산
해양과 대기 사이의 CO2 교환 플럭스(F, mmol C m−2 d−1)는 다음 식으로 결정된다(Takahashi et al., 2002).
여기서 k (m d−1)는 수온, 염분, 풍속으로 계산된 CO2의 기체 교환 속도이며(Wanninkhof, 2014), α (mol L−1 atm−1)는 수온과 염분으로 계산된 CO2의 용해도이다(Weiss, 1974). fCO2sw – fCO2atm (µatm)는 해양과 대기의 CO2 분압 차이이다. 이 식에 따라서 플럭스는 해양과 대기의 분압 차이에 비례하며, 음수의 플럭스는 대기에서 해양으로의 CO2 흡수를 의미한다.
2.2 18O 배양법을 이용한 유광대 총일차생산과 순군집생산 추정
2.2.1 시료 채수와 배양
배양 전 후 용존 O2의 동위원소 비 변화에 근거하는 18O in vitro 방법(Bender et al., 1987; Ferrón et al., 2016)으로 울릉분지와 경주 인근 연안 정점에서 유광대의 총일차생산(gross primary production; GPP)과 순군집생산을 추정하였다(Fig. 1의 St. 1과 St. 2). CTD rosette system에 부착된 PAR (photosynthetically active radiation) 센서를 이용하여 5개의 광 수심(표층 광량 대비 100, 50, 30, 12, 1%)을 결정하였다. Niskin 채수기의 해수를 산세척한 붕규산 유리병(140 mL)에 병 부피의 2 배 만큼 흘러 넘치게 한 후, H218O 시약(순도 97.2%; Sigma-Aldrich) 0.3 mL를 피펫 끝을 유리병 바닥에 가까이 넣어 주입하였다. 채수를 마친 유리병은 스크린(LEE Filters)을 붙여 광량을 조절한 용기에 넣은 후 표층수가 지속적으로 공급되는 배양 수조에 넣어 24시간 동안 배양하였다. 배양을 마친 시료는 산소 교환을 최소화하기 위해 질소 기체로 유리병 상부에 압력을 가해 시료를 2 개의 40 mL 유리병(serum bottle)에 나눠 담고 100% 염화 수은을 40 µL 주입한 후 밀봉하여 보관하였다. 용존 O2의 동위원소는 박막 주입식 질량분석기(membrane inlet mass spectrometer)를 이용하여 분석하였다(Kana et al., 1994; Ferrón et al., 2016). 분석 시 약 30분 마다 포화된 해수의 O2 동위원소 비를 측정하여 기기의 변동성을 관찰하였다. 중복 분석한 시료 간의 동위원소 비 차이는 ∼0.5‰이었다.
해수의 용존 O2 농도를 적정하기 위하여 Niskin 채수기의 해수를 마개가 있는 유리병(125 mL)에 병 부피의 2 배 만큼 흘러 넘치게 한 후, 중복으로 획득하였다. 분석은 하루 내에 선상에서 실시되었으며, 분석까지 대기와의 기체 교환을 최소화하기 위해 시료 병의 목 부분을 물로 채워 보관하였다. 용존 O2는 Winkler 적정법을 응용한 전류 측정기로 분석되었으며(Langdon, 2010), 중복 분석한 시료 간의 차이는 ∼0.3%였다. 적정법으로 결정한 산소 농도로 CTD rosette system에 부착된 O2 센서(Sea-Bird, SBE 43)를 보정하고, St.1 과 St.2의 광 수심에서 적정한 O2 농도는 다음 절에서 기술하는 총일차생산과 순군집생산 추정에 사용하였다.
2.2.2 총일차생산과 순군집생산 계산
질량분석기를 이용하여 산소의 무거운 동위원소(18O16O)와 가장 많이 존재하는 가벼운 동위원소(16O16O) 사이의 비를 측정하고, 이 둘의 동위원소 비(18R)를 계산한다. 포화된 해수의 O2 동위원소 비를 이용하여 기기 변동을 보정하였다(Kroopnick and Craig , 1972). 총일차생산(GPP, µmol O2 L−1 d−1)은 배양 기간 동안 용존 O2의 동위원소 비 변화를 토대로 추정되며, 식은 아래와 같다(Bender et al., 1987, 1999).
여기서 18R(O2)initial과 18R(O2)final은 각각 배양 전 후 O2의 동위원소 비이다. 18R(H2O)는 시약의 주입으로 농축(enrichment)된 해수의 동위원소 비이며, 시료병의 부피와 주입된 H218O 시약의 양으로 계산한다. [O2]initial (µmol L−1)은 배양 전 광 수심의 O2 농도이다. 배양 기간 동안 순산소변화를 나타내는 O2/Ar의 변화를 함께 측정하여 아래의 식으로 순군집생산(NCPin vitro, µmol O2 L−1 d−1)도 추정한다(Bender et al., 1999).
여기서 ([O2]/[Ar])initial과 ([O2]/[Ar])final은 각각 배양 전후 O2와 Ar 농도 비이다. 식 (5)와 식 (6)의 차이를 이용하여 총일차생산과 순군집생산의 차이로 정의되는 군집 호흡(community respiration; CR)까지 계산한다. 배양은 24시간 동안 실시되었기 때문에 위 값들은 모두 유광대 내의 하룻동안 생산 혹은 호흡을 나타낸다.
3. 결 과
3.1 표층의 용존 기체 분포
2019년 6월 5일부터 10일까지 동해 남서부해역 표층에서 관측한 자료들의 시계열 변화와 공간 분포를 Figs. 2와 3에 나타내었다. 수온(SST)과 염분(SSS)의 분포는 울릉분지와 삼척 및 포항 연안에서 큰 차이를 보였다. 울릉분지의 SST와 SSS는 각각 18.8 ± 0.2°C와 34.46 ± 0.05로 상대적으로 고온 고염한 분포를 보였다. 삼척 연안의 SST와 SSS는 각각 13.3 ± 0.4°C와 33.96 ± 0.03, 포항 연안의 SST와 SSS는 각각 17.5 ± 0.6°C와 33.78 ± 0.02로 분지에 비해 저온 저염한 특성을 보였다(Figs. 2(a), (b), Table 1). 가장 저온한 해수는 10.7°C로 삼척 연안에서, 가장 저염한 해수는 33.62로 경주 연안의 St.2 근처에서 관측되었다(Figs. 2(a), (b)).
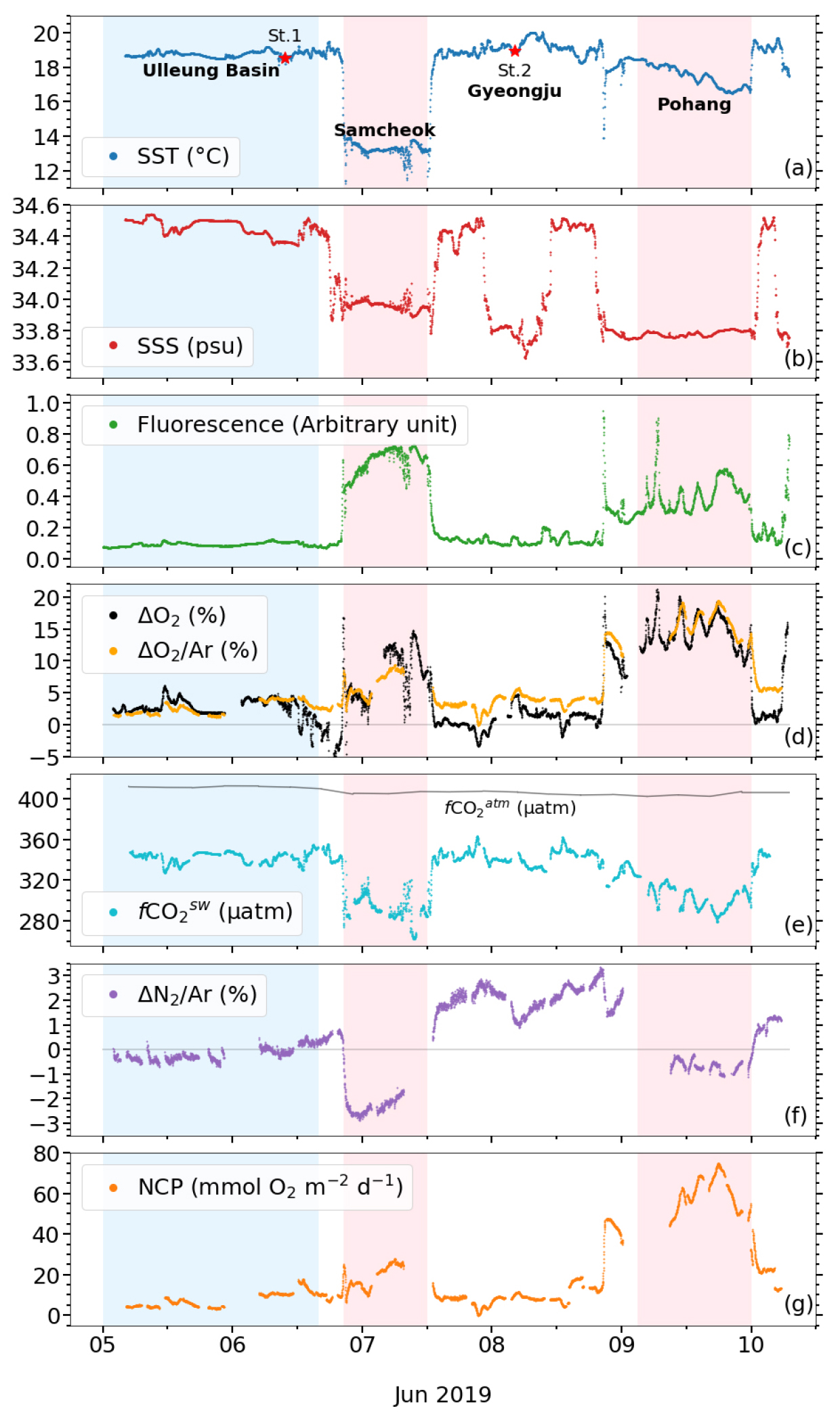
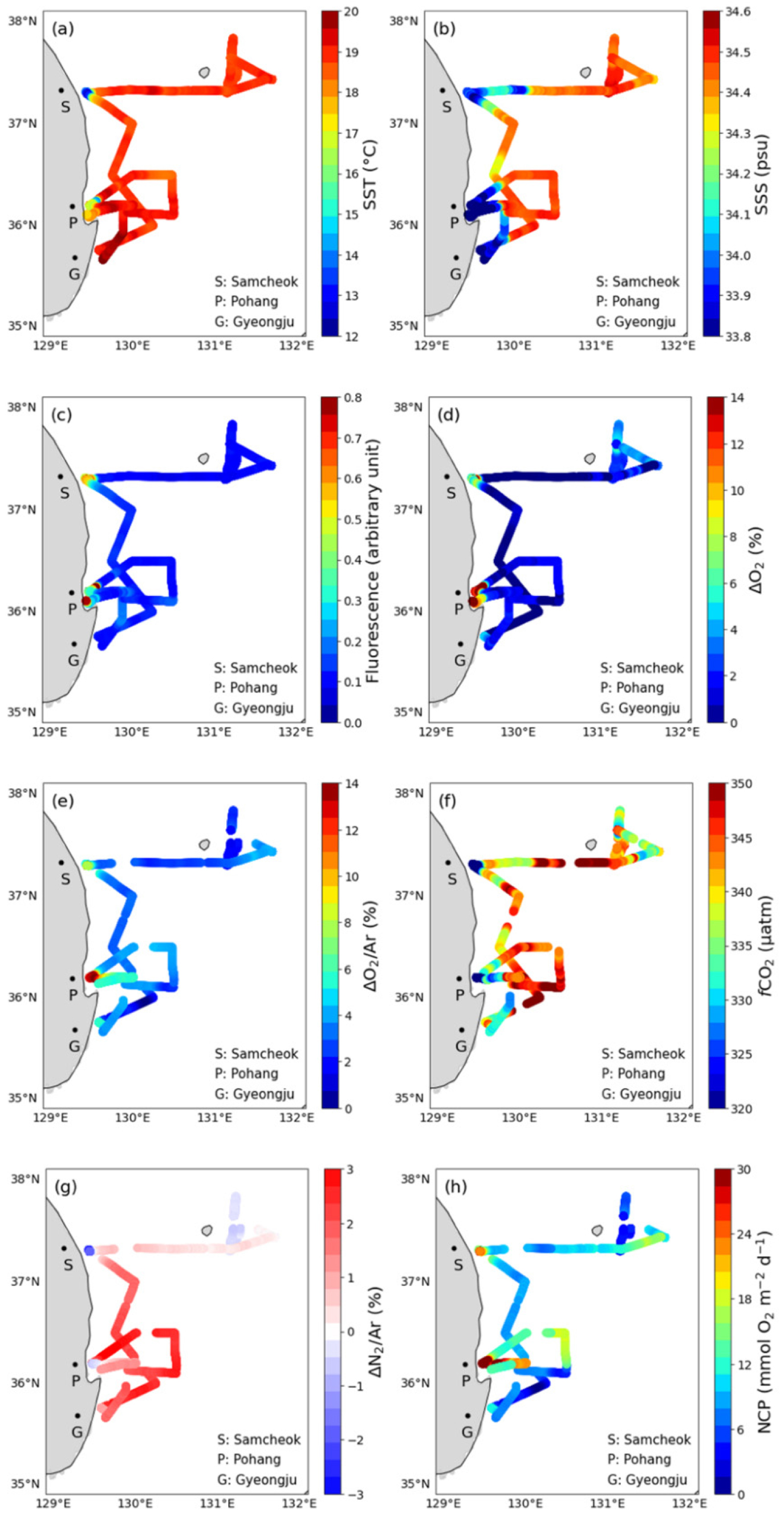
연안에서는 SST와 SSS 뿐만 아니라 용존 기체 용해도 특성에도 큰 변화가 나타났다. 식물플랑크톤의 생물량을 대변하는 엽록소 형광은 0.09 ± 0.01의 울릉분지에 비해 삼척과 포항 연안에서는 각각 0.63 ± 0.08과 0.41 ± 0.10로, 울릉분지에 비하여 약 4 배에서 7 배 가량 높았다(p < 0.05; Fig. 2(c), Table 1). ∆O2/Ar은 조사 해역에서 대부분 양의 값을 보이는데, 이는 관측 시기인 초여름 동해 남서부해역의 물질 대사가 독립 영양 환경임을 시사한다. 또한, ∆O2/Ar은 ∆O2(≡[O2]/[O2]sat − 1)와 변화 양상이 거의 동일한 것은 물론이고(Fig. 2(d)), 전 해역에서 평균 약 1.4%의 차이만을 보였다. 이는 동해 남서부해역의 O2 과포화가 대부분 생물학적으로 만들어졌음을 시사한다. 엽록소 형광의 변화와 마찬가지로, ∆O2/Ar의 분포는 울릉분지의 2.6 ± 1.0%인 것에 비하여 삼척과 포항 연안에서는 각각 6.4 ± 1.9%와 16.1 ± 2.0%로 분지에 비하여 약 3 배에서 6 배 가량 높았다(p < 0.05; Fig. 2(d), Table 1). 삼척과 포항 연안의 fCO2sw는 291 ± 12 µatm과 302 ± 11 µatm으로, 울릉분지의 341 ± 5 µatm에 비하여 각각 약 15%와 11% 만큼 낮았다(p < 0.05; Fig. 2(e), Table 1). fCO2atm은 전체 조사 기간의 평균이 408 ± 3 µatm으로 변동성이 작았다(Fig. 2(e)). 모든 해역에서 fCO2sw가 fCO2atm보다 낮아 해양이 대기 이산화탄소를 흡수하는 환경이었다. 위 기체 포화도 변화를 종합할 때, 삼척과 포항 연안에서는 활발한 생물 생산이 일어났던 것으로 생각된다.
∆N2/Ar은 연구선이 울릉분지에 머물던 6월 6일까지는 0%에 가까운 값을 보이다가 높은 풍속과 파도로 인해 연구선이 삼척 연안에 정박한 동안(6월 7일 오전)에는 최대 -3%까지 감소하였다(Figs. 2(f), 3(g)). 여기서 ∆N2/Ar은 ∆O2/Ar(식 (2) 참고)과 유사하게 다음과 같이 정의하였다.
∆N2/Ar은 삼척에서 포항으로 이동하는 구간에서는 2 – 3%로 증가하였다가 포항 인근 해역에서 -1%로 감소하였다(Figs. 2(f), 3(g)). 삼척과 포항 인근 해역 모두에서 ∆N2/Ar은 외해에 비해 약 3% 가량 감소한 것이다. 두 연안 해역에서 나타난 ∆N2/Ar의 감소가 엽록소 형광과 ∆O2/Ar의 증가, fCO2의 감소와 함께 나타나고 있어 이 해역의 용존 기체 용해도 변화가 일차 생산과 밀접한 관계가 있음을 시사한다(4.3 절 참고).
3.2 유광대 총일차생산과 순군집생산
울릉분지에 속하는 St.1과 경주 연안에 위치한 St.2에서 선상 배양을 이용해 유광대 내의 총일차생산(GPP), 순군집생산(NCPin vitro), 군집 호흡(CR)을 추정하였다(Fig. 4). St.1과 St.2에서 혼합층은 각각 12 m와 17 m에 형성되어 있었다(Figs. 4(a), (b)). 혼합층 깊이는 10 m 수온보다 0.2°C 낮은 수심으로 결정하였다(Lim et al., 2012). 유광대 내에서 St.1과 St.2의 수온 범위는 각각 12.2 – 18.5°C와 13.4 – 18.6°C로 유사하였으며, 혼합층 하부의 수온 구배 역시 두 정점에서 유의미한 차이가 나타나지 않았다(Fig. 4(a)). 이는 St.2가 삼척과 포항보다 남쪽 연안에 위치한 까닭에 고온한 대마난류수의 영향을 더욱 많이 받아 수온이 높아진 결과로 생각된다. 수온 분포와는 달리, 염분의 분포는 두 공간에서 차이를 보였다. St.1의 염분 범위는 34.31 – 34.37로 유광대 내에서 비교적 일정하였으나, St.2의 염분 범위는 33.74 – 34.36으로 혼합층 상부와 하부 사이의 구배가 컸다(Fig. 4(b)). 울릉분지와 연안 모두 유광대 깊이는 약 30 – 35 m 사이로 유사하였으나, 포항 연안의 아표층 엽록소 최대층(subsurface chlorophyll maximum layer; SCM)은 약 18 m로 유광대 내부에 존재한 반면, 울릉분지의 SCM은 약 38 m로 유광대 바로 아래에 위치하였다(Fig. 4(c)).
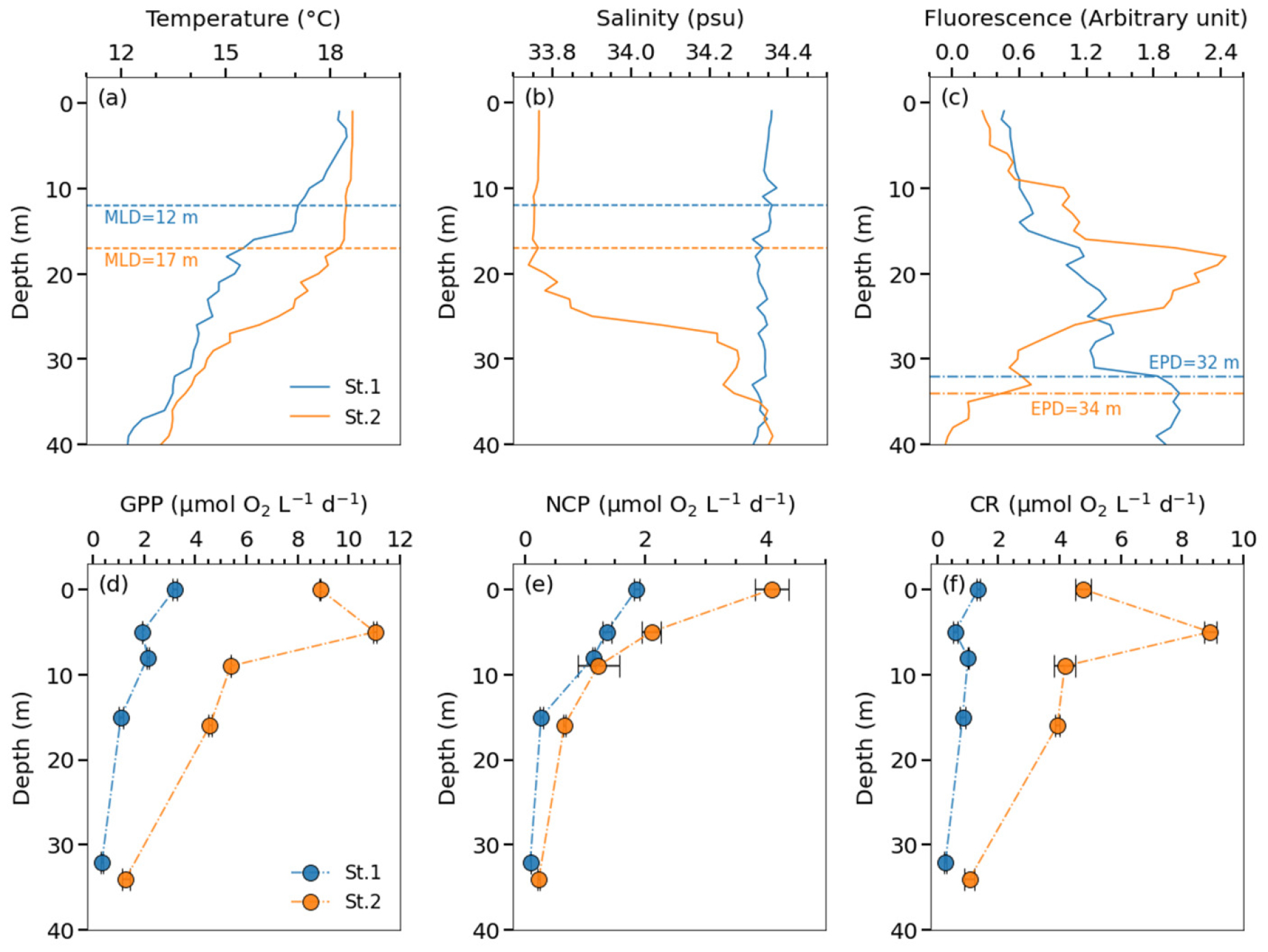
St.1과 St.2에서 GPP는 각각 0.4 – 3.2와 1.3 – 11.0 µmol O2 L−1 d−1의 범위를, NCPin vitro는 0.1 – 1.9와 0.2 – 4.1 µmol O2 L−1 d−1의 범위를, CR은 0.3 – 1.3과 1.1 – 8.9 µmol O2 L−1 d−1의 범위를 보였다(Figs. 4(d)~(f)). 표층 관측 결과와 마찬가지로, 유광대 내의 NCPin vitro는 모두 양의 값으로 나타나 수층이 독립 영양 상태임을 시사한다. GPP와 NCPin vitro는 수심에 따라 감소하는 PAR의 분포와 유사하게 표층에서 높고, 유광대 하부에서 낮은 양상을 보였다(Figs. 4(d), (e)). 전반적으로 수심별 NCPin vitro는 GPP에 비례하였으나, St.2의 50% 광수심에서는 표층에 비하여 GPP는 증가하였지만, NCPin vitro는 감소하였다(Figs. 4(d), (e)).
St.1과 St.2에서 깊이로 적분한 GPP는 각각 43과 170 mmol O2 m−2 d−1로 연안이 울릉분지보다 약 4 배 이상 높았다. 깊이로 적분한 NCPin vitro는 St.1과 St.2에서 각각 20과 37 mmol O2 m−2 d−1로 연안이 울릉분지보다 약 2 배 가까이 높았다. 1.4의 광합성 지수를 고려하였을 때(Laws, 1991), St.1과 St.2의 GPP는 각각 0.37과 1.46 g C m−2 d−1에 해당하며, NCPin vitro는 각각 0.17과 0.32 g C m−2 d−1에 상응한다. CR은 St.1과 St.2에서 각각 23과 134 mmol O2 m−2 d−1로 연안이 6 배 이상 더 높았다. 이는 연안이 많은 생물량으로 생물 생산이 더 활발하게 일어나지만, 군집의 호흡에 소모되는 유기물 역시 상당히 많은 환경임을 시사한다.
4. 토 의
4.1 연안과 울릉분지의 생산력 차이
울릉분지와 연안의 생산력을 비교하기 위하여, 생물 활동을 지시하는 항목들을 정점별로 Fig. 5에 나타내었다. 빗금이 없는 막대는 배양 조사로 얻은 GPP, NCPin vitro, CR이며 빗금 친 막대는 표층 ∆O2/Ar을 기반으로 추정한 혼합층 내의 NCPin situ이다. 배양 조사로 얻은 항목들은 유광대와 혼합층을 기준으로 각각 적분되었다. ∆O2/Ar을 기반으로 추정한 NCPin situ는 삼척과 포항 연안에서 각각 19 ± 6과 60 ± 9 mmol O2 m−2 d−1로 울릉분지의 8 ± 4 mmol O2 m−2 d−1에 비하여 약 2 – 7 배 가량 더 높았다(p < 0.05; Fig. 5(a), Table 1). 광합성 지수를 1.4 (Laws, 1991)로 가정하면, 삼척과 포항 연안의 평균 NCPin situ는 0.16 과 0.51 g C m−2 d−1에, 울릉분지의 평균 NCPin situ는 0.07 g C m−2 d−1에 상응한다. 경주 연안 St.2의 NCPin situ는 10 ± 1 mmol O2 m−2 d−1 (=0.09 g C m−2 d−1)로 울릉분지와 유사한 수준이었다(Fig. 5(a)). 이는 울릉분지와 비교하여 생물 펌프가 활발히 작동하는 두 연안과는 다른 양상이었다. St.2는 연안에 위치하여 염분이 낮다는 점만 제외한다면, 수온과 용존 기체 포화도 변화가 울릉분지와 유사하여(Fig. 2) 삼척 및 포항 연안과는 그 환경이 구분되는 것으로 생각된다.
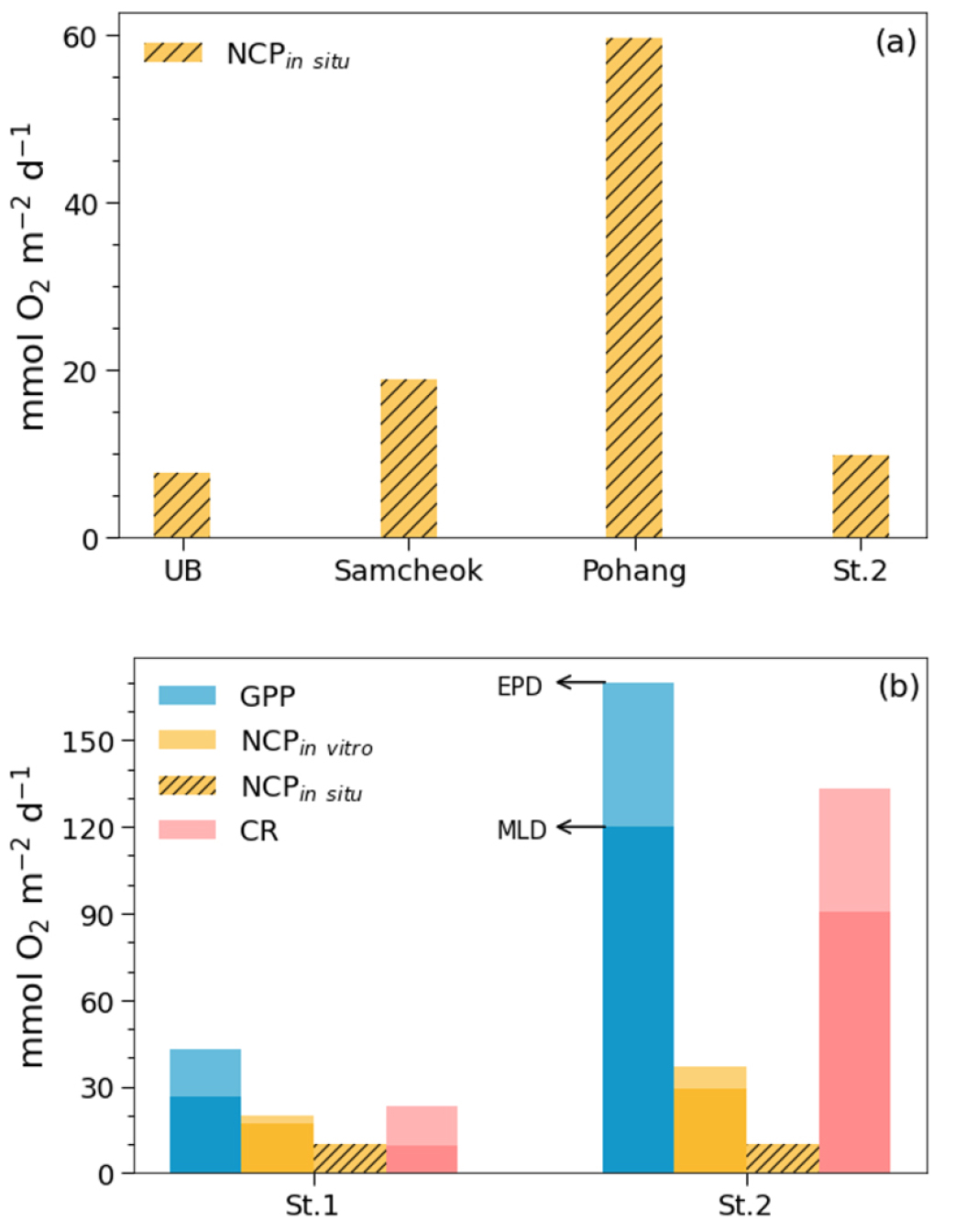
Fig. 5.
(a) The estimates of net community production in the mixed layers (NCPin situ) at Samcheok, Pohang, St.2, and in the Ulleung Basin. (b) Gross primary production, net community production (NCPin vitro), and community respiration determined by 18O in vitro method at St.1 and St. 2. NCPin situ is also shown for comparison with NCPin vitro.
생물 펌프의 관점에서 유사한 특성을 가지는 울릉분지와 경주 연안에 속하는 St.1과 St.2의 유광대 생산력을 비교하였다. 혼합층 순군집생산을 나타내는 NCPin situ와 NCPin vitro의 용이한 비교를 위해, 그리고 혼합층 하부 유광대의 생산력을 살펴보기 위해 18O in vitro 방법으로 측정한 수심별 생산력을 혼합층과 유광대 두 수심에 대해 적분하여 제시하였다(Fig. 5(b)). 유광대까지 적분한 GPP와 NCPin vitro에서 혼합층의 GPP와 NCPin vitro가 차지하는 비율이 약 60 – 85%로, 두 정점 모두 혼합층 내부에서 대부분의 일차 생산이 일어났다(Fig. 5(b)). 이 같은 결과는 여름철 울릉분지와 용승의 영향이 없는 연안에서 유광대까지 적분된 NPP 중 혼합층까지 적분된 NPP가 차지하는 비율이 약 20%에 불과한 연구 결과와는 차이가 있는 것이다(Kwak et al., 2013). Kwak et al.(2013)의 조사는 8월에 수행된 것으로 혼합층이 약 5 m에 형성되어 약 15 m에 형성된 본 연구의 혼합층보다 얕다. 제한된 정점이지만 두 연구 간의 조사 시기와 혼합층 차이를 감안할 때, 본 조사 시기에서는 성층이 채 강하게 발달되지 않아 빛 조건이 더 우세한 혼합층 내에서 많은 일차 생산이 일어난 것으로 생각된다.
동해에서 동일한 방법으로 GPP와 NCPin vitro를 추정한 연구가 없어, 13C와 15N 방법으로 추정한 NPP와 NP를 본 연구에서 추정한 값들과 비교하였다. Kwak et al.(2013)이 보고한 NPP의 범위는 울릉분지에서 0.37 – 0.96 g C m−2 d−1 (n=5)이었고, 연안에서는 0.45 – 2.41 g C m−2 d−1 (n=5)였다. 광합성 지수 1.4 (O2:C; Laws, 1991)를 가정하면, 본 연구에서 추정한 GPP는 St.1과 St.2에서 0.37과 1.46 g C m−2 d−1로, Kwak et al.(2013)이 보고한 NPP 범위에 속하였다. GPP가 NPP와 독립영양생물의 호흡에 소모된 것까지 모두 합한 양이긴 하나, 방법의 차이 뿐만 아니라 제한된 정점 수로 인해 두 생산력의 직접적인 비교는 어려웠다.
Redfield ratio를 고려하면(C:N=106:16) Kwak et al.(2013)이 보고한 NP의 범위는 울릉분지에서 0.05 – 0.40 g C m−2 d−1 (n=5)이었고, 연안에서는 0.14 – 1.91 g C m−2 d−1 (n=5)였다. NP는 유광대 하부에서 내부로 공급되어 이용 가능한 질소(주로 질산염)에 의해 유지되는 일차 생산으로, 정상 상태일 경우 NCP와 동일한 것으로 간주된다. 본 연구에서 추정한 NCPin vitro는 St.1과 St.2에서 0.17과 0.32 g C m−2 d−1로, 방법의 차이에서 나오는 오차를 고려한다면 Kwak et al.(2013)의 NP 추정치와 유사하였다.
GPP에 대한 NCPin vitro의 비(NCPin vitro/GPP)는 유광대 내에서 만들어진 유기 탄소 중에서 심층으로 수송되는 비율을 의미한다. 이는 여러 해역의 탄소 수송 효율을 비교하기에 적합한 지표이다. St.1과 St.2에서 혼합층 깊이까지 적분된 NCPin vitro/GPP는 0.65와 0.24이고, 유광대 깊이까지 적분된 NCPin vitro/GPP는 0.46과 0.22이었다. 이는 경주 연안이 울릉분지에 비하여 절대적인 생산력은 크지만, 많은 양의 유기물이 분해되어 탄소 수송 효율은 울릉분지와 유사함을 의미한다.
본 연구에서 독립적인 두 방법으로 NCP를 추정한 결과, 방법 간에는 약 2 – 3 배의 차이가 존재하였다(Fig. 5(b)). in situ 방법은 혼합층 내 O2의 체류 시간인 2 – 3주 간의 생산력을 반영(Teeter et al., 2018)하는 반면, in vitro 방법은 배양한 하루 간의 생산력을 반영한다. 이 점에서 방법 간의 차이가 비롯될 수 있긴 하나, 일반화를 하기에는 각 해역을 대표하는 정점이 하나씩 밖에 없어 어려움이 따른다. 향후 해역의 물질 대사에 대한 종합적인 평가를 위하여, 지속적인 현장 관측으로 각 방법을 이해하는 것이 필요하다.
4.2 연안과 울릉분지의 CO2 흡수력
해양과 대기 사이의 fCO2 차이로 계산된 CO2 교환율은 울릉분지에서 -4.7 ± 2.5 mmol C m−2 d−1이었고, 삼척과 포항 연안에서는 각각 -17.1 ± 8.9와 -25.8 ± 13.2 mmol C m−2 d−1이었다(Table 1). 이는 삼척과 포항의 단위 면적당 CO2 흡수율이 울릉분지에 비하여 약 4 배에서 5 배 가량 높음을 시사한다(p < 0.05). St.2의 CO2 흡수율은 -3.62 ± 0.75 mmol C m−2 d−1로 울릉분지와 유사한 수준이었다. 동해 남서부해역은 봄에 CO2를 흡수하고, 여름에 CO2를 방출한다고 알려져있다(Kim et al., 2014). 본 연구 시기 동안은 전 해역에서 CO2의 흡수가 일어나므로 봄철과 유사한 환경인 것으로 생각된다. 실제로 본 연구에서 관측한 울릉분지의 해양 대기 사이 fCO2 차이는 -71 µatm으로, Kim et al.(2014)이 보고한 5 – 6월의 관측값(-51 ± 20 µatm)과 유사하다.
3.1 절의 표층 연속 관측 결과는 울릉분지에 비하여 삼척과 포항에서 엽록소 형광, ∆O2/Ar 및 NCPin situ가 높게 나타나 연안에서 더 많은 일차 생산이 일어났음을 시사한다. 높은 일차생산력은 용존 CO2 흡수를 촉진하여 fCO2를 낮출 수 있다. 그러나 삼척, 포항 및 St.2 연안에서는 수온과 염분에도 큰 변 화가 있어(Figs. 2(a), (b)), 연안의 높은 CO2 흡수 요인을 살펴보기 위해서는 수온과 염분이 다른 해수의 혼합에 따른 fCO2sw 변화를 함께 고려해야 한다. 여름철 동해 연안에 영향을 줄 수 있는 해수는 고온 고염한 외해수, 러시아 혹은 북한 연안에서 생성되어 연안을 따라 남하하는 북한냉수(Yun et al., 2004)를 우선 꼽을 수 있다. 여기에 더해 삼척 연안에서 관측된 낮은 염분을 설명하기 위해 강을 통한 담수의 공급을 가정할 수 있겠다. 이 세 단성분 중 외해수의 수온과 염분은 울릉분지 해역의 관측값 평균(각각 18.8°C와 34.46; Table 1)으로 하였다. 북한냉수의 수온과 염분은 Yun et al.(2004)이 제시한 기준(염분 34.1 미만, σθ 27 이상)과 2001년 겨울 일본분지 관측 자료(KH42 cruise of CREAMS; http://sam.ucsd.edu/jes_atlas/.index.htm)를 근거로 1°C와 34.03으로 하였다. 담수의 수온과 염분은 16°C(관측 시기 기온)과 0으로 정했다. 세 해수의 혼합으로 연안의 수온과 염분이 결정되었다고 가정하면, 외해수, 북한냉수, 담수가 삼척 연안에서는 68%, 31%, 1% 비율로, 포항 연안 에서는 91%, 7%, 2% 비율로, 경주 인근 St.2 연안에서는 96%, 2%, 2% 비율로 혼합된 것으로 추정되었다.
Table 1.
Water properties (average ± standard deviation) of Ulleung Basin, Samcheok, and Pohang
해수 혼합으로 인한 fCO2sw의 변화를 추정하기 위하여, 해양에서 보존적인 추적자로 활용될 수 있는 총알칼리도(total alkalinity; TA)와 용존 무기 탄소(dissolved inorganic carbon; DIC)를 이용하였다(Dinauer and Mucci , 2018). 외해수, 북한냉수, 담수의 TA와 DIC 초기 농도를 각각 2284과 2025 µmol kg −1 (Kim et al., 2019), 2280과 2199 µmol kg −1 (KH42 cruise), 1705와 1735 µmol kg −1 (Xiong et al., 2019)이라 가정하였다. 위 혼합 비율로 계산한 TA와 DIC 농도는 삼척 연안에서는 2277과 2076 µmol kg −1, 포항 연안에서는 2273과 2032 µmol kg −1, St.2 연안에서는 2273과 2022 µmol kg −1로 추정되었다. 이 TA와 DIC 농도로 CO2SYS (Pierrot et al., 2006)를 이용하여 계산된 fCO2sw는 세 연안 순서로 392, 372, 368 µatm이었다(Fig. 6). 해수 혼합만으로 계산된 값(Fig. 6의 빈 동그라미)과 관측한 값(Fig. 6의 채워진 동그라미)의 차이는 생물학적 CO2 흡수에 해당하는 것으로 삼척 연안에서는 68 – 131 µatm, 포항 연안에서는 49 – 95 µatm, St.2 연안에서는 23 – 24 µatm 범위로 나타났다. 위 계산에서 가정한 북한냉수 단성분 특성의 불확실성을 고려하면, CO2 흡수력을 정교하게 평가하기 위해서는 추후 현장 관측 자료들을 보완할 필요가 있다.
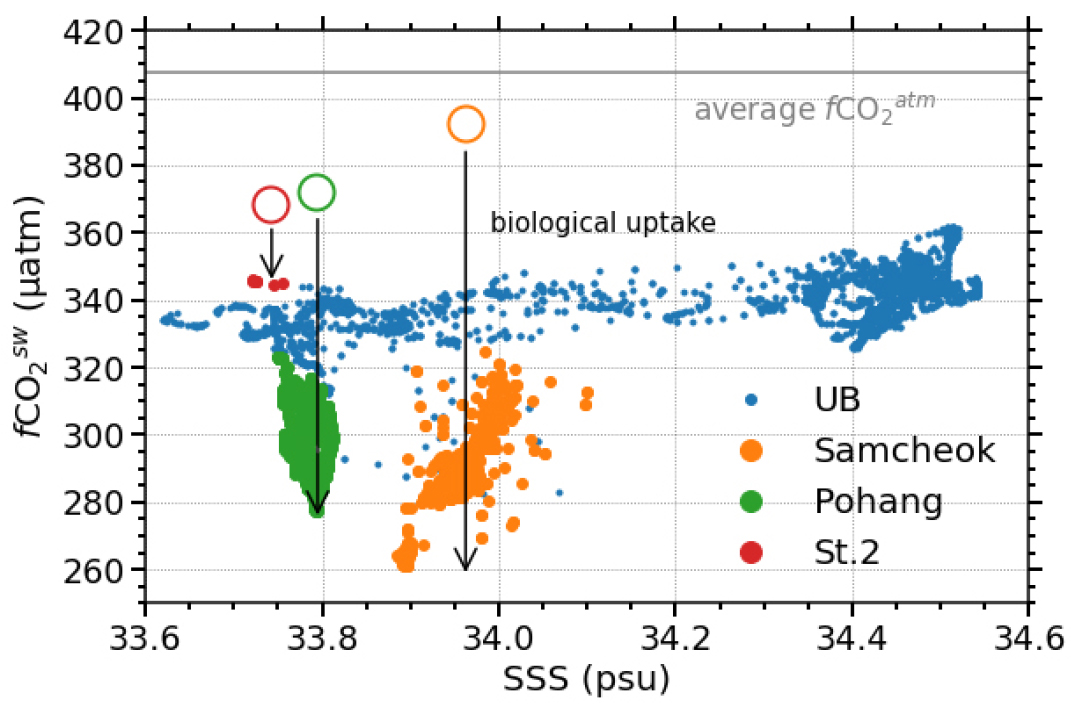
fCO2sw를 조절할 수 있는 또 다른 인자인 기체 교환과 해수 혼합 등의 영향은 생물 활동에 비하여 미미한 수준이었다. 혼합층 내 CO2의 체류 시간이 길어 해양과 대기 사이 기체 교환은 천천히 일어나기 때문에 이 영향은 제한적일 것이다(Choi et al., 2021). 또한, 분지와 연안의 수온 및 염분 프로파일은 수층 내에 성층이 발달되어 수직 혼합의 영향이 미미할 것을 암시하며(Figs. 4(a), (b)), 수평 이류의 영향은 대부분 위에서 가정한 해수 혼합에 반영되었을 것으로 생각된다. 4.1 절과 본 절의 내용을 종합했을 때, 삼척과 포항 연안에서 나타난 높은 NCPin situ는 연안에서 생물 펌프가 활발하게 작동하고 있음을 시사하고, 이 생물 펌프가 연안의 CO2 흡수를 촉진해 fCO2 분포를 조절한 것으로 여겨진다.
4.3 연안의 ∆N2/Ar 변화와 질소 고정 가능성
Ar을 포함한 비활성기체들은 화학 반응성이 없어서 이 기체들의 농도는 수온에 따른 용해도 변화, 기압 변화, 공기 주입(air injection)과 같은 물리적 요인에 의해서만 결정된다. N2의 수온에 따른 용해도 변화 정도는 Ar과 유사하여 N2/Ar비는 수온과 기압 변화에 의해서는 거의 변화하지 않는다(Hamme and Emerson, 2013). 반면, 공기 주입은 N2/Ar비가 84 (=0.7808/0.00934; Glueckauf, 1951)인 기체를 38인 해수(20°C, 염분 35 기준)에 추가하는 과정으로 ∆N2/Ar을 증가시킨다. 해양 표층의 기체들은 확산에 의한 대기-해양 기체 교환과 공기 주입(air injection)의 경쟁 결과로 대체로 포화 혹은 약간 과포화(1 – 3%) 상태에 있다(Hamme and Emerson, 2002).
6월 8일 전후 관측 해역의 ∆N2/Ar이 2 – 3%로 높아진 것은 높은 파도로 인해 공기 주입 효과가 강화된 결과로 여겨진다(Fig. 2(f)). 비교적 파도가 낮았던 시기(6월 7일 12:00시 전)에는 평균 6 m/s의 바람이 불었으나, 이후 연구선이 피항을 위해 삼척 연안에 머무르던 6월 8일 00:00시까지는 8 – 14 m/s의 강한 바람이 불었다. 이 때 공기 주입에 의한 과포화가 8일 삼척에서 포항 인근 해역으로 이동하는 중에 관측이 된 것으로 여겨진다. 약 0.6 mL의 공기가 포화 상태의 해수 1 kg에 주입되면 ∆N2/Ar은 2.5%로 나타날 수 있다(Pilson, 2013). 6월 6일과 8일 관측 해역의 수온 변화는 미미하여 8일 ∆N2/Ar이 증가한 현상을 설명할 수 없다. 또, 기압에 따른 두 기체의 포화도 변화 정도는 동일하여 ∆N2/Ar에는 변화가 없다(Hamme and Emerson, 2013).
삼척과 포항 연안(6월 7일과 9일)에서는 ∆N2/Ar이 외해(6월 6일과 8일)에 비해 최대 3% 감소하였다(Fig. 2(f)). 수온과 염분 변화에 의한 ∆N2/Ar 변화 정도를 살펴보기 위해, 4.2 절에서 제시한 외해수, 북한냉수, 담수의 혼합 비율과 각 해수에 N2와 Ar이 포화되어 있다는 가정을 이용하여 혼합 해수의 ∆N2/Ar을 계산하였다(Hamme and Emerson, 2004). 계산된 ∆N2/Ar은 삼척과 포항 연안에서 각각 -0.28%와 -0.12%로, 세 해수 혼합에 따른 수온과 염분 변화는 삼척과 포항 연안의 ∆N2/Ar 감소의 극히 일부만 설명할 수 있다. 앞서 언급한 것처럼 기압 감소는 두 기체 포화도 변화에 동일하게 작용하여 ∆N2/Ar 에는 변화가 없다. 요약하면, 삼척과 포항 연안의 ∆N2/Ar 감소를 설명할만한 물리 요인이 존재하지 않는다.
EIMS를 이용한 한 연구에서는 Baltic Sea에서 -8 – 3%의 ∆N2/Ar을 관측하였고, 음과 양의 포화도를 각각 질소 고정과 공기 주입의 결과로 해석하였다(Schmale et al., 2019). 삼척과 포항 인근 해역의 ∆N2/Ar 감소가 ∆O2/Ar의 증가와 fCO2sw의 감소를 수반하고 있다는 점은 일차 생산자에 의한 질소 고정이 활발히 일어났을 가능성을 시사한다. 최근 동해 아한대 해역에서 질소 고정 군집에 관한 연구가 보고되었다(Sato et al., 2021). ∆O2/Ar의 기체 교환으로부터 NCP를 추정한 것과 같은 방식으로(식 (3) 참고), ∆N2/Ar의 기체 교환으로부터 질소 고정률(nitrogen fixation; NF)을 다음과 같이 계산하였다.
N2의 기체 교환 속도 kN2와 포화 농도 [N2]sat을 각각 1.0 m d−1 , 468 µmol kg−1으로 하고, ρ와 ∆N2/Ar을 각각 1025 kg m−3과 2%(삼척 연안 평균)로 가정하면 NF는 19 mmol m−2 d−1에 이른다. 이 값은 Tang et al.(2019)이 그동안 보고된 질소 고정률의 상위 2%에 해당한다고 언급한 New Jersey 연안의 3 mmol m−2 d−1보다도 6 배 가량 큰 것으로, 동해 남서부해역 연안에서도 질소 고정이 매우 활발히 일어나고 있음을 의미한다. 앞에서 살펴본 것처럼 공기 주입에 의한 ∆N2/Ar 편향이 양의 방향으로 일어남을 고려하면, ∆N2/Ar을 이용한 이 추정은 질소 고정률의 하한선에 해당한다. N:O2를 16:138으로 가정하면, 삼척 연안의 NF는 NCP의 8 배에 해당하는 것이다. 향후 보다 정교한 측정법(예를들면, Klawonn et al., 2015; Cassar et al., 2018)을 이용하여 동해 남서부해역의 질소 고정률을 살펴볼 필요가 있겠다.
5. 결 론
그동안 많은 연구가 수행되지 않았던 동해 연안의 생물 펌프 및 탄소 흡수력을 이해하기 위하여, 삼척과 포항 연안의 표층 ∆O2/Ar 및 fCO2 변화를 울릉분지와 비교하였다. 삼척과 포항 연안의 NCPin situ는 각각 19 ± 6과 60 ± 9 mmol O2 m−2 d−1로 울릉분지의 8 ± 4 mmol O2 m−2 d−1에 비하여 약 2 – 7 배 가량 높았다. 연안에서 활발하게 작동하는 이 생물 펌프는 해양의 fCO2 감소와 그에 따른 대기 CO2 흡수로 이어졌다. 생물학적인 CO2 흡수가 삼척과 포항 연안에서 -17.1 ± 8.9, -25.8 ± 13.2 mmol C m−2 d−1로 울릉분지에 비해 4 – 5 배 가량 높았다. 또한 두 연안에서는 ∆N2/Ar이 최대 3% 감소하였는데, 생물 생산력이 높은 이 해역에서 질소 고정이 일어났을 가능성을 시사한다.
경주 연안의 NCPin situ는 10 ± 1 mmol O2 m−2 d−1로, 다른 두 연안과 달리 울릉분지와 생물 펌프 세기에는 큰 차이가 없는 것으로 나타났다. 이처럼 연안 내에서도 생물 펌프 세기에는 공간 변동성이 존재한다. 또한 본 연구는 6월에 제한되어 계절 변화를 포함한 연안의 탄소 흡수 특성을 이해하기에는 자료가 부족하다. 향후 지속적으로 현장 관측을 수행한다면 탄소 순환에서 동해 연안이 차지하는 중요성을 이해할 수 있을 것이다.
