

1. 서 론
2. 재료 및 방법
2.1 실험대상종
2.2 실험방법
2.3 통계분석
3. 결 과
3.1 수온에 따른 포식성 기생생물의 발달과 세대시간의 변화
3.2 수온에 따른 포식성 기생생물의 생존율과 감염성공률의 변화
4. 토 의
1. 서 론
무각 와편모류 Akashiwo sanguinea는 광온성 및 광염성 종으로 전 세계 기수역과 연안환경에서 연중 빈번하게 출현하는 종으로 잘 알려져 있다. 이 종에 의한 적조는 1922년 일본 내만에서 최초로 보고된 이후(Hirasaka 1922), 미국(Martin 1929; Cardwell et al., 1979; Bockstahler and Coats, 1993; Jessup et al., 2009), 멕시코(Kiefer and Laasker, 1975), 캐나다(Robinson and Brown, 1983), 호주(Hallegraeff 1991), 중국(Lu and Hodgkiss, 2004), 흑해(Gomez and Boicenco, 2004), 한국(Lee et al., 2005), 북해(Gomez and Souissi, 2008) 등을 비롯하여 전 세계 연안에서 흔히 발생하는 것으로 보고되고 있다. 이 종에 의한 적조 피해는 국내에서는 아직 보고된 적이 없지만, 페루 연안과 미국 캘리포니아 Monterey 만에서 적조가 발생했을 때 어류와 바다새가 집단 폐사하였다고 보고하였다(Jessup et al. 2009). 또한 일본에서는 A. sanguinea에 의한 적조가 김의 황백화 현상을 유발하는 원인으로 보고하였으며, 뿐만 아니라 전복의 유생을 폐사시키거나, 굴의 성체와 유생에 급성 독성을 나타내어 수산양식산업에 큰 경제적 손실과 피해를 유발할 수 있는 것으로 보고하였다(Cardwell et al., 1979; Botes et al., 2003; Matsubara et al., 2007).
일반적으로 A. sanguinea의 출현 시기는 우리나라 남해안의 경우, 2월에서 11월에 걸쳐 연중 빈번하게 출현하면서, 주로 고수온기인 5~9월에 출현밀도가 상대적으로 높은 것으로 보고되었다(Lee et al. 2005). 그러나 최근 거제도 연안에서 2011년 저수온기인 추계-동계(11~1월)에 매우 고밀도(~107cells/L)의 A. sanguinea에 의한 적조를 형성하여 약 2개월에 걸쳐 장기간동안 지속되는 이상 현상이 보고되었다(Son and Baek, 2012). 일부 연구에서는 A. sanguinea에 의한 적조 발생의 주요 요인으로서 영양염 이용능력, 광범위한 수온과 염분에 대한 내성, 다른 생물을 섭식하는 혼합 영양학적 특성 등을 비롯한 다양한 환경적 요인들이 작용한다고 보고하였다(Nakamura et al., 1982; Bockstahler and Coats, 1993; Robinson and Brown, 1983).
와편모류에 의한 적조를 제어하는 주요 기작으로서 동물플랑크톤에 의한 섭식이나 포식성 기생생물에 의한 감염 등의 생물학적 요인에 의한 하향 조절(top-down control)에 의한 제어를 들 수 있다(Chambouvet et al., 2008; Mazzillo et al., 2011; Kim et al., 2013; Worden et al., 2015). 그중에서 포식성 기생생물 Amoebophrya에 의한 감염은 적조의 급격한 쇠퇴 현상과 밀접한 관련이 있다는 연구 결과가 발표되었다(Taylor et al., 1968; Nishitani et al., 1984; Coats et al., 1996; Chambouvet et al., 2008; Kim and Park, 2014). 포식 기생(parasitoidism)이란 숙주를 감염시킨 후 숙주물질을 이용하여 단시간(약 2일 이내)안에 성장하여 기생생물의 성숙이 완료되면, 숙주를 죽이고 수많은 감염성 쌍편모자들을 배출해 내는 특성을 가진다. 프랑스 Penzé 하구역에서 3년 동안 와편모류에 의한 적조와 포식성 기생생물 Amoebophrya의 개체군의 거동을 조사한 결과, 다른 여러 환경요인보다 와편모류의 개체수 변화 및 종천이 양상이 숙주 특이성이 강한 포식성 기생생물 Amoebophrya에 의한 감염과 매우 밀접한 연관성을 가지는 것으로 보고하였다(Chambouvet et al., 2008). 이러한 포식 기생생물이 적조생물을 제어할 수 있는 능력은 감염성 쌍편모자 한 개체가 하나의 적조생물 세포를 감염시킨 후, 단시간(약 2일)안에 102~103개 이상의 새로운 세대의 감염성 쌍편모자들을 생산하여 해수 중으로 방출되기 때문에 숙주인 적조생물의 성장률보다 훨씬 더 빠른 증식속도와 높은 생산력을 나타낼 뿐 아니라, 적조생물을 섭식하는 다른 포식자의 섭식률이나 번식률과 비교해도 월등히 높기 때문이다. 일부 연구에서는 포식성 기생생물 Amoebophrya의 증식률과 숙주 감염률, 총 세대시간 등의 변수들을 이용하여 수치 모델링을 이용하여 분석한 결과, Amoebophrya에 의한 감염이 다른 포식자(미소동물플랑크톤)의 섭식과 같은 생물학적 조절 요인보다 적조를 제어하는데 훨씬 더 큰 영향을 미칠 수 있다고 발표하였다(Montagnes et al., 2008).
포식기생성 와편모류 Amoeobophrya spp.에 의해 감염된 숙주 군집은 현재 적조를 유발하는 종들을 포함하여 약 75종의 다양한 와편모류 종들을 감염시키는 것으로 알려져 있다(Kim et al., 2004; Park et al., 2013; Kim and Park, 2014). 최근 차세대 염기서열 분석방법을 이용하여 빈영양인 지중해를 비롯한 대서양과 태평양, 남극해 등 전 세계 다양한 해역에서 조사한 결과, 전체 원생생물 군집에서 매우 높은 비율을 차지할 뿐 아니라, 기존에 알려지지 않은 매우 높은 종다양성이 규명되어 해양 생태계에서 이들의 생태적 기능과 역할에 대해 현재 큰 주목을 받고 있다(Guillou et al., 2008; Cleary and Durbin, 2016; De Vargas et al., 2015). 적조생물을 감염시키는 포식성 기생생물의 감염력에 영향을 미치는 환경적 요인으로는 영양염에 대한 연구가 유일하며(Yih and Coats 2000), 수온과 같은 물리적 환경요인에 대한 연구는 거의 전무하다. 따라서 본 연구에서는 유해 적조를 유발하는 와편모류 A. sanguinea와 이를 감염시키는 포식성 기생생물 Amoebophrya sp.의 숙주-기생생물 시스템을 이용하여 수온에 따른 포식성 기생생물의 감염성공률과 생존율의 변화에 대해 알아보았고, 각 수온에 따라 포식성 기생생물의 감염율과 숙주 세포내 발달 및 총 세대시간의 변화를 측정하였으며, 이러한 결과를 바탕으로 실제 해양생태계에서 숙주개체군 제거율에 미치는 영향에 대해 규명하고자 하였다.
2. 재료 및 방법
2.1 실험대상종
본 연구에 이용된 유해 적조 유발성 와편모류 Akashiwo sanguinea와 이를 감염시키는 포식성 기생생물 Amoebophrya sp.의 숙주-기생생물 시스템은 미국 스미스소니언 환경연구소에서 분양받아 이용하였다(Coats and Park, 2002). 이 숙주-기생생물 배양체는 염분 15에서 분리 배양되었으나, 본 실험에 이용하기 위해 염분을 서서히 증가하여 최종 29에 적응시켜 이용하였다. 숙주인 A. sanguina는 염분이 29인 멸균 여과해수에 규산염을 제외한 f/2-Si 배지(Guillard and Ryther, 1962)를 이용하여 배양하였다. 포식성 기생생물 Amoebophrya sp.는 2-3일 간격으로 주 3회마다 지수성장기의 숙주를 공급하는 계대배양으로 유지하였다. 배양의 온도와 염분은 각각 20oC와 29에서 수행하였고, 광도는 80 µmol m-2s-1(형광램프)의 14L:10D의 광주기 조건에서 배양하여 실험에 이용하였다.
2.2 실험방법
2.2.1 수온에 따른 포식성 기생생물의 발달과 세대시간의 변화
숙주는 지수성장기의 A. sanguinea 배양체를 각 3개 반복구의 500 mL 플라스크에 300 mL씩 나누어 담은 후 각 플라스크를 초기 배양온도인 20°C에서 각 실험 온도(15°C, 20°C, 25°C)에 적응시키기 위해 매일 1°C씩 서서히 증감하여 각 실험온도에 도달하도록 하였다. 최종적으로 각 온도에서 적응시킨 배양체를 100 mL씩 분주하여 3개의 반복구를 설정하여 실험에 이용하였다. 포식성 기생생물 배양체도 숙주와 같은 방법으로 각 온도 조건에서 숙주 배양체와 함께 서서히 적응시켜 실험에 이용하였다. 숙주의 초기 세포밀도는 약 1000 cells/mL이 되도록 접종하였고, 포식 기생생물은 숙주에서 빠져 나온 지 6시간 이내의 쌍편모자(dinospore)를 수확하여, 숙주와 포식성 기생생물의 개체수 비율이 1:5가 되도록 접종하여 실험을 진행하였다. 매 6-8 시간 간격으로 피펫을 이용하여 일정량의 시료를 분취하여 최종농도 1%가 되도록 글루타르알데하이드를 첨가하여 고정하였다. 고정한 시료는 Sedgwick Rafter 계수기를 이용하여 Coats and Bockstahler (1994)의 방법에 따라 청색 필터(Filter set 09, BP 450-490 excitation, beam splitter FT 510, emission LP515)가 장착된 형광현미경(Axio Imager A2, Carl Zeiss, Germany)하에서 감염된 숙주 세포수를 계수하여 감염율을 측정하였고, 이 때 포식성 기생생물의 세포밀도는 혈구계수기(hemocytometer)을 이용하여 계수하였다. 포식 기생생물의 숙주 세포내 발달시간은 Coats and Bockstahler (1994)의 방법에 의해 계산하였고, 포식 기생생물의 총 세대시간(Generation Time; GT)은 Coats and Park (2002)에 따라 기생생물 쌍편모자를 숙주에 접종하여 숙주를 감염시킨 다음, 숙주를 죽이고 다시 새로운 세대의 쌍편모자가 출현하기 시작하여 최대 세포밀도를 나타내는 시점까지의 세포밀도를 각 시간에 따라 면적을 적분한 다음, 적분한 면적의 1/2에 해당하는 시간을 계산하여 산출하였다.
2.2.2 수온에 따른 포식성 기생생물의 생존율과 감염성공률의 변화
수온에 따른 포식 기생생물의 생존율과 감염성공률의 변화를 측정하기 위해, 위에서 기술한 바와 같이 각 실험온도(15°C, 20°C, 25°C)에서 서서히 적응시킨 숙주-기생생물 배양체를 이용하였다. 각 온도 조건하에서 포식 기생생물의 생존율을 알기 위해, 감염된 숙주세포에서 빠져나온 지 6시간 이내의 기생생물의 쌍편모자를 멸균된 여과지(10 µm pore size, polycarbonate filter)를 이용하여 중력식 여과법(gravity filtration)으로 걸러서 숙주 세포를 제거한 다음, 각 실험온도에서 시간에 따라 쌍편모자의 세포밀도의 변화를 측정하였다. 이는 일정시간 간격에 따라 피펫을 이용하여 3 ml의 시료를 취한 후, 최종농도 1%가 되도록 글루타르알데하이드를 첨가하여 고정한 다음, 형광현미경하에서 혈구계수기(hemocytometer)를 이용하여 계수하였다. 각 온도별 쌍편모자의 시간에 따른 사멸속도(k)는 쌍편모자 세포밀도 변화 자료를 다음과 같은 단일이변량지수감소곡선(single two-parameter exponential decay curve)식에 대입하여 비선형 최소자승법으로 각 변수들을 구하였다.
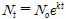
t: 시간(h)
Nt: t시간 경과 후 세포밀도(cells/mL)
No: 초기 세포밀도(cells/mL)
k: 세포의 사멸속도(mortality rate)
각 온도별 시간에 따른 포식성 기생생물의 숙주세포에 대한 감염성공률(successful infection rate of dinospores, %)은 숙주세포에 첨가한 기생생물 쌍편모자 개체수에 대한 성공적으로 숙주를 감염시킨 쌍편모자 개체수의 비율을 말하며, 이를 측정하기 위해 기생생물의 쌍편모자를 수확하여 각 실험 온도(15°C, 20°C, 25°C)에서 일정시간 간격에 따라 쌍편모자를 숙주인 A.sanguinea (1000 cells/ml)에 1:1(v/v) 비율로 접종시켜 36시간동안 배양한 다음, 형광 현미경하에서 성공적으로 숙주를 감염시킨 쌍편모자를 계수하여 Coats and Park(2002)의 방법에 따라 감염성공률을 계산하였다.
2.3 통계분석
본 연구에서 측정한 자료는 평균(±표준오차)로 제시하였으며, 평균 간의 유의성을 검증하기 위해 IBM SPSS statistics 23.0 program을 이용하여 T-검정 및 일원분산분석(one-way ANOVA test)을 수행하여 분석하였다.
3. 결 과
3.1 수온에 따른 포식성 기생생물의 발달과 세대시간의 변화
포식성 기생생물 Amoebophrya의 쌍편모자(dinospore)를 숙주인 적조생물 A. sanguinea 배양체에 접종한 이후, 시간이 경과함에 따라 적조생물 세포내로 침투하여 감염시킨 후 숙주 물질을 이용하여 성장하면서 성숙이 완료되면 기생생물이 숙주를 죽이고 수중으로 빠져 나오면서 연충형(vermiform) 단계를 거쳐 다음 세대의 수백 개의 쌍편모자를 생성하였다(Fig. 1). 각 온도 조건에서 포식성 기생생물의 초기 감염율을 측정한 결과, 25°C와 20°C에서는 각각 53±2.2% (n=3, T40)와 38±0.4% (n=3, T60)를 나타낸 반면, 15°C에서는 28±0.8% (n=3, T87)를 보여 비교적 낮은 감염율을 나타내었다(Table 1). 포식 기생생물 Amoebophrya 쌍편모자의 세포 밀도는 숙주 A. sanguinea를 감염시킨 다음 성숙이 완료되어 숙주세포를 죽이고 다시 수중으로 방출되면서 시간에 따라 점차 급격하게 증가하는 경향을 보이며, 25°C와 20°C의 온도 조건에서는 접종한 지 64시간과 96시간 경과한 후에 각각 120±0.3×103cell/ml (mean±SE, n=3)와 125±0.7×103cell/ml (mean±SE, n=3)의 세포밀도를 나타내었고 다시 새로운 숙주세포를 감염시키면서 개체수가 감소하다가 다음 세대의 쌍편모자가 출현하는 경향을 나타내었다(Fig. 2(a),(b)). 반면에, 15°C에서는 이보다 상당한 시간이 더 소요되어 131시간이 경과한 후에 쌍편모자의 세포밀도가 30±0.7×103 cells/ml (mean±SE, n=3)에 도달하였다(Fig. 2(c)).

Fig. 1. Epifluorescence micrographs of Amoebophrya sp. and its host Akashiwo sanguinea. Free-living stage of the parasite dinospore(A), early infection where the green fluorescing parasite is located inside the nucleus of the host A. sanguineaand surrounded by red fluorescing chloroplasts of the host (B), and mid to late infection where the developed parasite trophont is occupied in the protoplasm of the host(C). Scale bars represent 20µm.
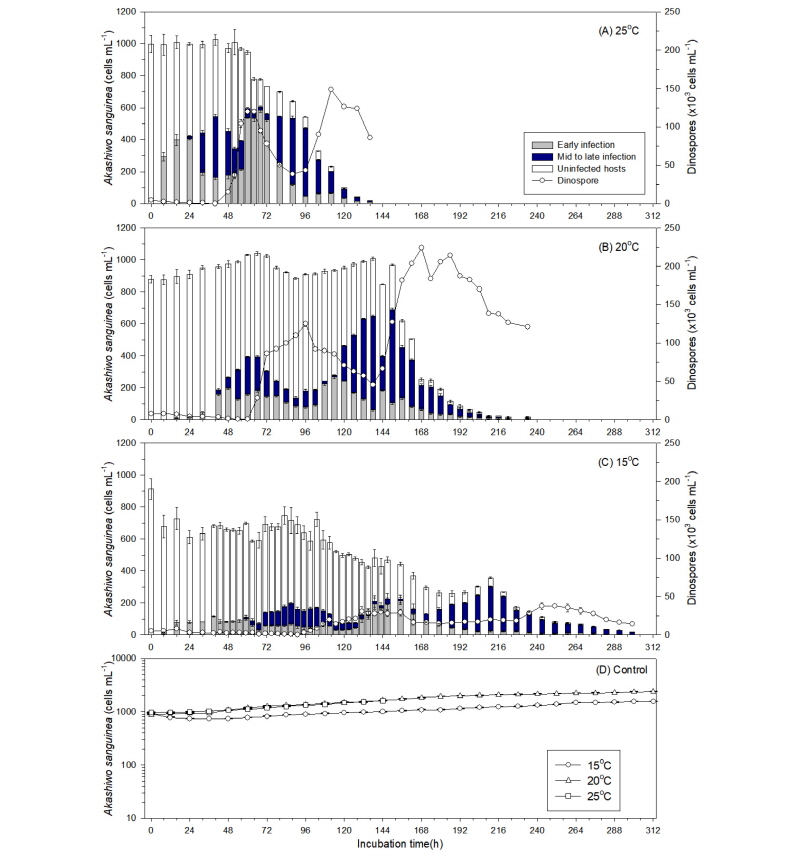
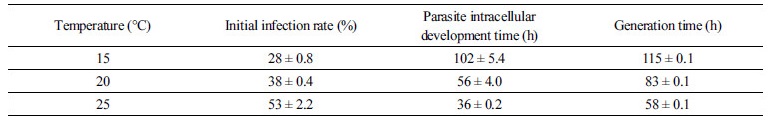
각 온도 조건 별 포식 기생생물의 숙주 세포내 발달 시간을 산출해 본 결과, 15°C, 20°C, 25°C에서 각각 102±5.4h, 56±4.0h, 36±0.2h의 범위로서 온도가 낮을수록 세포내 발달 시간이 매우 길어지는 것으로 나타났다(Table 1, One way ANOVA; post-hoc Tukey test; p<0.0001). 포식성 기생생물의 총 세대시간을 추정한 결과, 25°C에서는 약 58±0.1h과 20°C에서는 83±0.1h, 15°C에서는 115±0.1h의 범위를 나타내어 각 온도 조건에서 총 세대시간은 유의한 차이를 보였으며, 온도가 낮아질수록 포식성 기생생물의 세대시간이 급격하게 증가하는 것으로 나타났다(Table 1, one-way ANOVA; Tukey test; p<0.0001). 각 온도 조건에서 적조생물 A. sanguinea는 포식 기생생물의 감염과 발달이 진행되면서 25°C에서는 136시간, 20°C에서는 234시간, 15°C에서는 291시간이 경과 후에 모두 사멸하였다(Fig. 2). 반면에, 기생생물을 첨가하지 않은 대조구에서는 A. sanguinea의 세포밀도가 시간에 따라 점차 증가하여 실험기간동안 일정하게 유지되는 양상을 나타내었다(Fig. 2(d)).
3.2 수온에 따른 포식성 기생생물의 생존율과 감염성공률의 변화
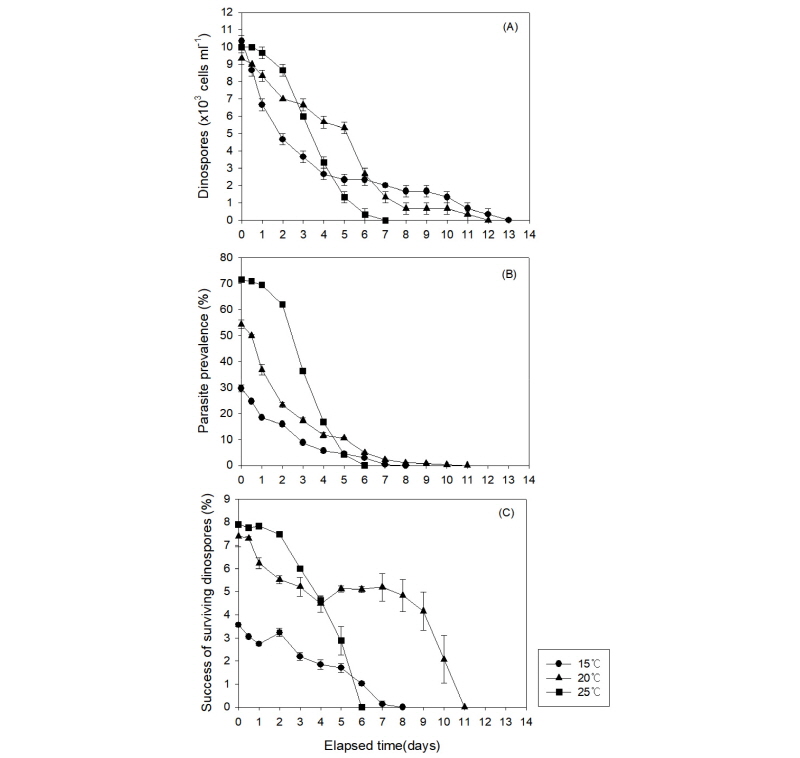
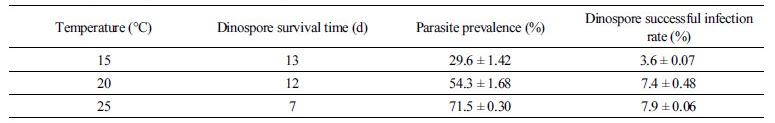
포식 기생생물 Amoebophrya의 쌍편모자는 숙주가 없는 조건에서, 시간이 지남에 따라 개체수가 점차 감소하는 양상을 나타내었으며, 25°C 온도에서 쌍편모자의 생존기간(survival time)은 7일로 나타났으며, 20°C에서는 12일, 15°C에서는 13일 경과 후에 모두 사멸하는 것으로 나타났다(Fig. 3(a)). 각 온도조건에서 시간에 따른 쌍편모자 개체수의 변화율을 단일이변량지수감소곡선식(single two-parameter exponential decay curve)에 대입하여 비선형 최소자승법으로 각 변수들을 구한 결과, 25°C에서 배양한 쌍편모류의 개체수는 시간에 따라 가장 높은 감소율(k=-0.30, r2=0.89, p=0.0015)을 나타내었고, 20°C(k=-0.20, r2=0.93, p<0.0001)와 15oC(k=-0.28, r2=0.96, p<0.0001)에서는 상대적으로 시간에 따라 낮은 개체수의 감소율을 나타내었다(Table 2, Fig. 3(a)). 15°C에서는 다른 높은 온도 조건보다 초기에 급격한 개체수의 감소를 나타내었지만, 시간이 지날수록 생존기간은 오히려 가장 긴 것으로 나타났다(Fig. 3(a)). 각 온도조건에서 감염율(parasite prevalence)은 25°C에서 71.5±0.30%의 범위로 가장 높게 나타났고, 20°C와 15°C에서는 각각 54.3±1.68%와 29.6±1.42%의 범위를 나타내어 온도가 낮아질수록 감염율이 크게 감소하는 것으로 나타났다(Table 3, Fig. 3(b), one-way ANOVA; Tukey test; p < 0.0001). 각 온도 조건에서 시간에 따른 숙주 A. sanguinea에 대한 쌍편모자의 감염성공율을 측정해 본 결과, 25°C와 20°C에서는 쌍편모자의 초기 감염성공율(To)이 각각 7.9±0.06%와 7.4±0.48%의 범위를 나타내었고, 15°C에서는 3.6±0.07%의 범위로 가장 낮은 수치를 나타내었다(Table 3, Fig. 3(c)). 그러나 시간이 경과할수록 25°C에서는 쌍편모자의 생존기간이 가장 낮았고, 감염률도 급격하게 낮아져 쌍편모자 개체당 감염성공률이 가장 빠르게 감소하여 6일 경과 시에 모두 감염능력을 잃은 것으로 나타났다(Table 3, Fig. 3). 반면, 15oC에서는 쌍편모자의 숙주생물에 대한 감염율이 시간에 따라 점차 감소하였지만, 생존기간은 가장 길게 나타나 8일 동안 감염능력을 유지하는 것으로 나타났다. 20°C에서는 쌍편모자의 생존기간이 길고 감염율이 느리게 감소하여 쌍편모자 개체당 감염성공률이 비교적 오랫동안 지속되면서 11일 동안 감염능력을 가지는 것으로 나타났다(Table 3, Fig. 3).

4. 토 의
포식 기생성 와편모류인 Amoebophrya는 유해 및 유독성 적조를 유발하는 다양한 와편모류를 감염시키며, 적조생물에 대한 생물학적 제어 요인으로 잘 알려져 있다(Talyor et al., 1968; Chambouvet et al., 2008). 유독 와편모류 Alexandrium minutum에 의한 적조가 매년 빈번하게 발생하는 프랑스 Penze 하구역에서 연구한 결과에 따르면 포식 기생성 와편모류 Amoebophrya spp. 감염이 다양한 와편모류에 의해 발생한 적조의 소멸과 천이 양상과 밀접한 연관성이 있음을 보고하였다(Chambouvet et al., 2008). 국내에서는 대표적으로 매년 유해 적조를 유발하는 Cochlodinium polykrikos를 비롯하여 유독 와편모류 Alexandrium spp. 혼합종과 Gonyaulax polygramma 및 Prorocentrum minimum 등의 적조 발생시 Amoebophrya에 의한 감염이 보고되었다(Kim et al., 2002; Kim et al., 2004; Park et al., 2007; Kim and Park, 2014). 해양생태계에서 포식성 기생생물 Amoebophrya의 감염으로 인한 와편모류 숙주개체군의 제거율(parasite-induced mortality)을 유추해 보았을 때, 약 ~30%day-1로 추정되었다(Coats and Bockstahler, 1994; Velo-Suάrez et al., 2013). 해양생태계에서 이러한 포식기생생물에 의한 숙주개체군의 제거율은 현장에서 출현한 적조생물에 대한 감염율이 높을수록 증가하는 반면, 기생생물의 숙주 세포내 발달시간 및 세대시간이 길어질수록 감소한다고 할 수 있다. 이러한 두 변수, 감염율과 세대시간은 다양한 물리화학적 환경요인에 의해 영향을 받을 수 있다. 한 예로 숙주세포의 영양학적 질과 밀접한 관련을 가질 수 있는 서식환경의 영양염 조건에 의해 영향을 받을 수 있다. 즉, 부영양 환경보다 빈영양 환경에서 성장한 숙주를 감염시키는 포식성 기생생물은 세대시간에서는 유의한 차이를 나타내지 않았지만, 숙주개체군에 대한 감염율은 빈영양환경보다 부영양 환경에서 더 높은 감염율을 나타내었다.(Yih and Coats, 2000). 그러나 지금까지 이러한 적조생물을 감염시키는 포식성 기생생물의 감염력에 대해 수온과 같은 물리적 환경요인이 미치는 영향에 대해 연구한 자료는 전무하다. 본 연구에서는 포식성 기생생물의 감염율과 세대시간에 대해 물리적 환경 요인인 수온에 따른 영향을 조사한 결과, 수온에 따라서 포식성 기생생물의 감염력이 유의한 차이를 나타낼 뿐 아니라, 포식성 기생생물의 숙주 세포내에서의 발달시간과 세대시간에서도 유의한 차이를 보이는 것으로 나타났다. 즉, 낮은 수온에서는 숙주 세포내 발달이 느려지게 되어 총 세대시간이 매우 길어지는 결과를 나타내었다. 이는 저수온기에 대증식한 와편모류 개체군에 대해 포식성 기생생물에 의한 감염율이 매우 낮아질 뿐 아니라, 감염이 되더라도 숙주내 포식성 기생생물의 발달이 매우 느리게 진행되어 총세대시간이 길어지게 되므로, 이는 결과적으로 해양 생태계내에서 숙주개체군 제거율이 매우 감소하게 된다. 따라서, 수온이 낮은 시기에 발생한 와편모류에 의한 동계 적조는 포식성 기생생물의 감염과 같은 하향 조절(top-down control)에 의한 영향을 상대적으로 적게 받을 것으로 예상된다. 다른 한편, 이는 숙주개체군의 밀도가 낮은 동계에 낮은 감염율과 숙주세포 내에서 포식성 기생생물의 발달시간이 느려짐으로써, 숙주개체군 제거 속도가 느려지고, 해양생태계 내에서 이용가능한 숙주개체군을 모두 사멸시키는 것을 막아 월동을 위한 하나의 생존전략이 될 수 있다. 즉, 본 연구 결과에서 나타난 것처럼 수온이 낮을수록 포식성 기생생물의 감염율은 상대적으로 낮아지지만, 포식성 기생생물의 개체수와 감염성공율이 상대적으로 천천히 감소하게 되어 감염력이 보다 오랫동안 유지되기 때문에 숙주 밀도가 낮은 동계시기동안 생존하는 데 유리할 것으로 판단된다.
적조 유발성 와편모류 A. sanguinea는 수온 16~22°C와 염분 22~34의 비교적 넓은 범위에서 출현하는 광온성 및 광염성 종으로 알려져 있다(Lee et al., 2005). 우리나라 남해안에서 조사한 1990~2003년까지의 자료에서는 A. sanguinea가 2월-12월까지 거의 연중 내내 출현하며, 주로 고수온기인 5월-8월에 상대적으로 높은 밀도(~10000 cells/L)로 출현한다고 보고되었다(Lee et al., 2005). 그러나 최근의 연구결과에서는 고수온기가 아닌 저수온기에서 A. sanguinea의 적조가 발생하였으며, 출현밀도도 고수온기보다 현저히 높은 고밀도로 한 달 이상동안 장기간 지속되었다고 보고하고 있다. 예를 들면, 2011년에 국내 진해만 부근에서 저수온기인 11월-12월에 이례적인 고밀도의 A.sanguinea의 적조(~107cells/l)가 발생하였다고 보고하였으며(Son and Baek, 2012), 이후 최근 2016년 남해안에서는 저수온기(13.8~17.3°C)인 11월-12월에 적조(100~1500 cells/ml)가 발생하였다(NFRDI, 2017). 이러한 저수온기에 발생한 고밀도의 적조 현상은 미국 캘리포니아 Monterey만에서도 보고된 바 있다(Mazzillo et al., 2011). 이에 따르면, 2005년-2007년 동안 3년 동안 현장 조사한 자료로서, 2005년과 2006년과는 달리 2007년 저수온기에 비교적 높은 밀도의 A. sanguinea의 적조가 발생하여 한 달 이상동안 장기간 지속되었다고 보고하였다. 적조발생시 표층수온은 다른 조사시기보다 낮았고, Amoebophrya에 의해 감염된 세포도 관찰되지 않았다고 보고하였다(Mazzillo et al., 2011). 이는 본 연구 결과에서 알 수 있듯이 20°C 이하로 수온이 낮아질수록 와편모류 개체군에 대한 감염율이 급격히 저하될 뿐 아니라, 감염 후 포식 기생생물이 숙주 세포내 성장 및 발달시간이 매우 길어지므로 감염능력을 가진 다음 세대의 포식성 기생생물 쌍편모자가 생성되는 데 걸리는 총 세대시간이 급격히 길어지게 된다. 결과적으로, 수온이 낮을수록 포식 기생생물에 의한 숙주개체군의 제거율을 감소시켜 저수온기에 A. sanguinea에 의한 고밀도의 적조가 형성되어 장기간 유지되었을 것으로 유추할 수 있다. 따라서 저수온기에는 숙주개체군에 대한 포식성 기생생물의 감염과 같은 하향조절에 의한 영향이 크게 낮아질 수 있으며, 이는 결과적으로 추계 및 동계 적조의 고밀도화와 장기화를 유발하는 요인이 될 수 있음을 시사한다.
한편, 와편모류 A.sanguinea에 대한 포식 기생생물 Amoebophrya의 숙주개체군 제거율에 영향을 줄 수 있는 요인으로서, 강한 숙주특이성을 가지는 포식성 기생생물이 숙주 종주(strain)에 따라 감염율의 차이를 나타낼 가능성을 배제할 수 없다. 즉, 고수온기에 출현하는 A. sanguinea 종주와 저수온기에 출현하는 종주가 서로 다를 수 있으며, 이는 숙주 종주에 따라 Amoebophrya의 감염율이 차이를 나타낼 가능성이 있다. 최근 연구결과에 따르면, 중국 연안에서 2008년-2012년 5년간 출현한 A. sanguinea의 유전자형을 분석하여 연구한 결과, 서로 다른 유전자형(genotype)인 A타입과 B타입이 혼재하여 출현하였으며, 또한 서로 다른 계절적인 출현양상을 나타내었다고 보고하였다(Luo et al., 2017). 즉, 유전자형 A타입은 일년 내내 출현하다가, 수온이 낮은 동계와 춘계에 상대적으로 높은 출현밀도를 나타낸 반면에, 유전자형 B타입은 고수온기인 하계에만 높은 밀도로 출현하는 것으로 조사되었다. 이 두 가지 유전자형의 A. sanguinea 종주를 단일세포로 분리하여 실내 배양실험을 수행한 결과, 유전자형 A타입은 저수온에서 상대적으로 높은 생장률을 나타낸 반면, 유전자 B타입은 고수온에서 더 높은 생장률을 보여 현장 모니터링 결과를 뒷받침하는 결과를 보였다. 일반적으로 포식 기생생물 Amoebophrya spp.는 숙주특이성이 다르며, 감염시킬 수 있는 숙주 종의 범위에 있어서 차이를 나타내는 것으로 알려져 있다(Coats et al., 1996; Kim 2006). 이 가운데, A.sanguinea를 감염시키는 Amoebophrya는 비교적 강한 숙주특이성을 나타내는 것으로 알려져 있으며 (Coats and Bockstahler, 1994) 숙주인 A.sanguinea의 유전자 타입에 따라 감염율의 차이를 나타낼 가능성이 있다. 즉, 하계에 적조를 일으키는 유전자형 B타입의 A.sanguinea를 감염시키는 포식성 기생생물이 다른 유전자형의 A타입을 감염시키지 못하거나 혹은 상대적으로 낮은 감염율을 나타낼 가능성이 있다. 이러한 가능성은 추후에 국내에서 저수온기와 고수온기에 출현한 A.sanguinea의 유전자형과 출현현황을 조사하고, 각 시기별 출현한 숙주의 유전자형에 대한 포식성 기생생물의 감염력이 유의한 차이를 나타내는지에 대한 추가적인 연구가 필요하다.
