1. 서 론
규조류는 현미경의 발달과 함께 18세기 초에 최초로 그 존재가 관찰되었다(Round et al., 1990). 담수와 해수 환경, 기수역, 토양 등 다양한 서식지에서 발견되고 있으며 많은 학자들에 의해 그 분류와 생태가 연구되고 있다. 해양과 담수 생태계에서 중요한 1차 생산자로서(Licea et al., 2016), 부유성 규조류 뿐 아니라 저서규조류 역시 높은 생물량과 생물다양성을 보인다. 갯벌 조간대에서 생물막을 형성하여 저서 동물의 중요한 먹이원이 될 뿐 아니라(Mitbavkar and Anil, 2002) 해양에서의 규소 순환 조절에 큰 기여를 하고 있다(Yool and Tyrrell, 2003). 또한 환경변화에 매우 민감하게 반응하는 규조류의 특성을 활용하여 유용한 수질 지표로도 활용되고 있다(Underwood and Kromkamp, 1999; Park and Koh, 2012).
한국에서 규조류 연구는 1920년대에 Skvortzow(1929)가 수원의 서호에서 자생하는 마름풀로부터 규조류 1신종, 1신변종을 포함해 58종의 규조류를 보고하면서 시작되었고, 그 뒤로도 외국학자들에 의한 몇 편의 연구가 수행되었다(Skvortzow, 1932; Kokubo, 1940). 국내학자에 의한 최초의 규조류 연구는 박태수(박, 1956a; 1956b)에 의해 이루어졌으며, 이후 1960년대 후반부터는 규조류 연구자와 논문 편수 모두가 크게 증가하였다(최, 1967, 1972; 이 등, 1967; 최, 1969; 정 등, 1971; 유, 1973, 1974; 유와 이, 1976; 박, 1979; Yoo and Lee, 1979). 1980년대 이후부터는 환경과 규조류 사이의 관련성에 대한 연구(박, 1980, 1982; Cho, 1981; 김과 조, 1985; 김, 1989; 유, 1980; 윤 등, 1991) 역시 활발하게 진행되었다. 이러한 노력들의 결과로 현재까지 한국의 담수 및 해양환경에서 약 1680여종에 이르는 규조류가 보고되었다(이와 조, 2015). 그러나 최근 연구 결과(Park et al., 2014)에 따르면 한국 규조류의 다양성은 여전히 저평가되고 있으며 훨씬 더 많은 수의 규조류 종들이 신종 혹은 미기록종으로서 기재되어야 할 것으로 여겨진다. 특히 갯벌 등 해양저서환경의 경우 규조류의 종다양성이 높은 반면 종 수준의 명확한 동정이 어려워 속 수준의 동정 및 보고에 그치는 경우가 많았다.
Navicula는 해양저서환경에서 가장 우점하는 대표 분류군 중 하나로서, 속명이 의미하는 바와 같이 작은 배와 닮은 형태를 갖는다(Round et al., 1990). 종다양성이 매우 높아 1200개 이상의 종이 속해 있으며(Bourasi et al., 2018), 그만큼 분류적 연구의 필요성도 크다. 일례로서 새만금 갯벌에서 이루어진 연구(Oh and Koh, 1995)의 경우 총 371개의 출현종 중 Navicula 속의 종들은 79종으로서 전체의 20%를 차지하였는데, 이들 중 약 절반가량인 36종은 속 수준의 동정에 그쳤다. 또한 Park et al.(2016)은 기존에 한국 연안에서 Navicula scopulorum으로 보고되고 있던 종이 실제로는 Climaconeis속의 신종임을 밝혀 새로이 기재한 바 있다. 이에 대해서 저자는 유럽과 미국에서의 규조류 연구 결과를 무비판적으로 수용한 데 그 원인을 두고 있다. 규조류 연구의 역사가 깊고 활발한 외국의 경우에도 기존에 Navicula로 분류되던 종들에 기반하여 2000년 이후에 여러 새로운 속이 정립된 바 있기도 하다. 예를 들면 Envekadea (Gligora et al., 2009), Pseudofallacia (Liu et al., 2012), 그리고 Decussiphycus (Guiry and Gandhi, 2019) 등이 이에 해당된다.
본 연구에서는 Navicula 속의 분류학적 연구를 위해 황해 11개 정점에서 갯벌 퇴적물을 채집하여 출현 규조류 종들을 관찰하였다. 광학현미경 및 전자현미경 관찰에 기반하여 상세한 관찰을 수행하였으며, 외형적 유사성뿐 아니라 미세구조의 특징 역시 엄밀하게 고려하여 동정을 수행하였다. 그 결과 기존에 흔히 Navicula flanatica (Grunow, 1860)로 동정되어 왔던 종의 특징이 해당 종의 원전과 일치하지 않으며, 오히려 개각의 크기 및 점무늬열의 형태와 밀도에 있어 Navicula spartinetensis (Sullivan and Reimer, 1975)와 잘 일치함을 발견하였다. 따라서 미기록종으로서 해당 종의 형태적 특징을 자세히 기재하여 보고하고자 하며, 더불어 동북아시아에서의 Navicula spartinetensis 분포에 대한 정보를 제시하고자 한다.
2. 재료 및 방법
2.1 연구 지역 및 시료 채집
황해는 한반도와 중국대륙에 둘러싸인 평균수심 44 m의 반폐쇄성 만으로서 대륙붕이 잘 발달되어 있다(Koh and Khim, 2014). 또한 중국에서는 황하강과 양자강이, 한반도에서 한강, 영산강과 금강 같은 하천에서 퇴적물이 유입되어, 연안에서는 점토가 우세한 조간대가 분포한다(Choi, 1991; Oh and Kum, 2001). 규조류 관찰을 위한 표층 퇴적물 시료는 황해의 해안선을 따라 한국과 중국의 여러 갯벌에서 각각 채집하였다(Table 1). 한국에서는 2006년 10월 강화도에서 2곳, 2007년 7월 서산에서 1곳, 2018년 7월 부안에서 1곳으로 총 4곳에서 채집하였다. 중국에서는 2018년 6월과 7월 사이에 랴오둥반도에서 2곳, 보하이만에서 1곳, 산둥반도에서 4곳으로 총 7곳에서 표층퇴적물을 채집하였다. 퇴적물 시료는 조수가 저 수위일 때 스파툴라로 퇴적물의 표층을 0-0.5cm 긁어서 채집하였으며, 코니컬 튜브에 담아 현장에서 해수와 포르말린을 섞어 고정하였다. 고정된 시료는 실험실로 가져온 후 현미경 관찰을 위한 전처리 과정을 진행하였다.
Table 1.
Sampling sites in the Yellow sea
|
Sample
|
Country
|
Station
|
Latitude (N)
|
Longitude (E)
|
Date
|
|
KO-BO
|
Korea
|
Hwado-myeon, Ganghwa-gun, Incheon, Korea
|
37°36' N
|
126"31' E
|
06.10.27
|
|
KO-OJ
|
Korea
|
Oji-ri, Daesan-eup, Seosan-si, Chungcheongnam-do, Korea
|
36"57' N
|
126"22' E
|
07.07.03
|
|
KO-YC
|
Korea
|
Yeocha-ri, Hwado-myeon, Ganghwa-gun, Incheon, Korea
|
37"35' N
|
126"22' E
|
06.10.24
|
|
KO-GS
|
Korea
|
Upo-ri, Julpo-myeon, Buan-gun, Jeollabuk-do, Korea
|
35"34' N
|
126"39' E
|
18.07.14
|
|
CH-BZ
|
China
|
Haixing, Cangzhou, Hebei, China
|
38"15' N
|
117"51' E
|
18.07.05
|
|
CH-DD
|
China
|
Gushanzhen, Donggang, Dandong, Liaoning, China
|
39"50' N
|
123"39' E
|
18.07.02
|
|
CH-DL
|
China
|
Zhuanghe, Dalian, Liaoning, China
|
39"39' N
|
122"59' E
|
18.07.02
|
|
CH-QD1
|
China
|
Chengyang, Qingdao, Shandong, China
|
36"15' N
|
120"19' E
|
18.06.30
|
|
CH-QD2
|
China
|
Xihai'an, Qingdao, Shandong, China
|
35"46' N
|
119"55' E
|
18.06.29
|
|
CH-RZ1
|
China
|
Donggang, Rizhao, Shandong, China
|
35"17' N
|
119"26' E
|
18.06.27
|
|
CH-RZ2
|
China
|
Ganyu District, Lianyungang, Jiangsu, China
|
35"04' N
|
119"18' E
|
18.06.27
|
2.2 시료 처리 및 분석
채집한 퇴적물 시료의 전처리는 Park(2011)의 방법에 따라 처리하였다. 먼저, 순차적인 초음파 처리를 통해 퇴적물로부터 규조류를 분리하였다. 분리된 규조류 시료는 염산(HCl)과 과산화수소(H2O2)를 이용해 유기물과 실트 입자를 제거하였다. 유기물 제거가 끝난 규조류 시료와 Naphrax 레진을 사용해 영구 프레파라트를 제작하여 광학현미경 관찰에 사용하였고, 시료의 일부는 스터브(stub)에 얹어 잘 건조시킨 후 백금코팅하여 주사전자현미경 관찰에 사용하였다.
규조류의 관찰 및 사진촬영을 위해서는 디지털 카메라(OLYMPUS Tech X Cam Ⅲ, ZEISS Axiocam 305 color)가 부착된 광학 현미경(Olympus BX53, x100 대물렌즈)을 사용하였다. 또한 광학 현미경에서 관찰이 어려운 규조류의 미세구조 관찰 및 촬영을 위해서는 전계방사형 주사전자현미경(Field Emission Scanning Electron Microscope; Tescan, MIR-3)을 사용하였다. 마지막으로 촬영한 규조류 사진의 도판 편집에는 Adobe Photoshop elements 10 프로그램이 이용되었다.
3. 결과 및 고찰
Navicula flanatica는 한국 갯벌에서 흔히 보고되어 왔던 종으로서, 조(1989) 및 Park(2011) 연구 등에 도판이 제시된 바 있다. 본 연구에서 11개 정점 모두에서 동일한 종이 출현하였으나, 원전에 기재된 Navicula flanatica의 원래 형태적 특징과는 일치하지 않음을 확인하였다. 또한 기존의 Navicula 연구 문헌을 살펴 본 결과, 황해 지역에서 Navicula flanatica로 동정되어 보고된 종의 특징은 개각의 크기 및 점무늬열의 형태와 밀도에 있어 Navicula spartinetensis와 잘 일치하였다. 따라서 본 연구는 Navicula spartinetensis의 국내 출현을 보고하는 최초의 연구이다.
3.1 현미경 관찰 결과 및 분류학적 고찰
3.1.1 황해출현 Navicula spartinetensis의 형태적 특징
연구지역에서 출현한 종의 광학현미경과 주사전자현미경으로 관찰한 내용은 다음과 같다.
광학현미경 관찰 : 개각(valve)은 좁은 피침형(laceolate)으로서(Fig. 1), 끝(apex)쪽으로 향할수록 점점 가늘어진다. 개각의 윤곽은 조금씩 형태적 차이를 갖는다(Figs. 1(a-b), Figs. 1(d-e)). 개각의 길이는 19.3-29.3 ㎛이며, 폭은 4-6 ㎛이다. 점무늬열(stria)의 밀도는 10 ㎛에 13-15개이며, 그물눈(areola)의 밀도는 10 ㎛에 20개였다. 등줄(raphe)은 직선형으로서 단순한 구조를 갖는다. 등줄 가슴판(raphe sternum)은 넓고 두꺼우며(Figs. 1(c-d)), 중앙 영역(central area)에서 가로방향으로 약하게 확장한다. 중앙 영역의 등줄 끝(Central raphe ending)은 단순하고, 작은 점처럼 약간 확장된다(Fig. 1). 그리고 양쪽 등줄의 끝은 같은 방향으로 강하게 굽어 있다. 그물눈(areola)은 선형(lineolate)으로서 명확하게 관찰된다. 점무늬열은 중앙 영역에서는 방사형(radiate)이나, 양 끝 쪽 방향으로 갈수록 끝볼록마디(terminal nodule) 방향으로 수렴(convergent)한다(Figs. 1(b-c)). 점무늬열은 등줄 가슴판을 중심으로 약간 비대칭을 이룬다(Figs. 1(a-c)).
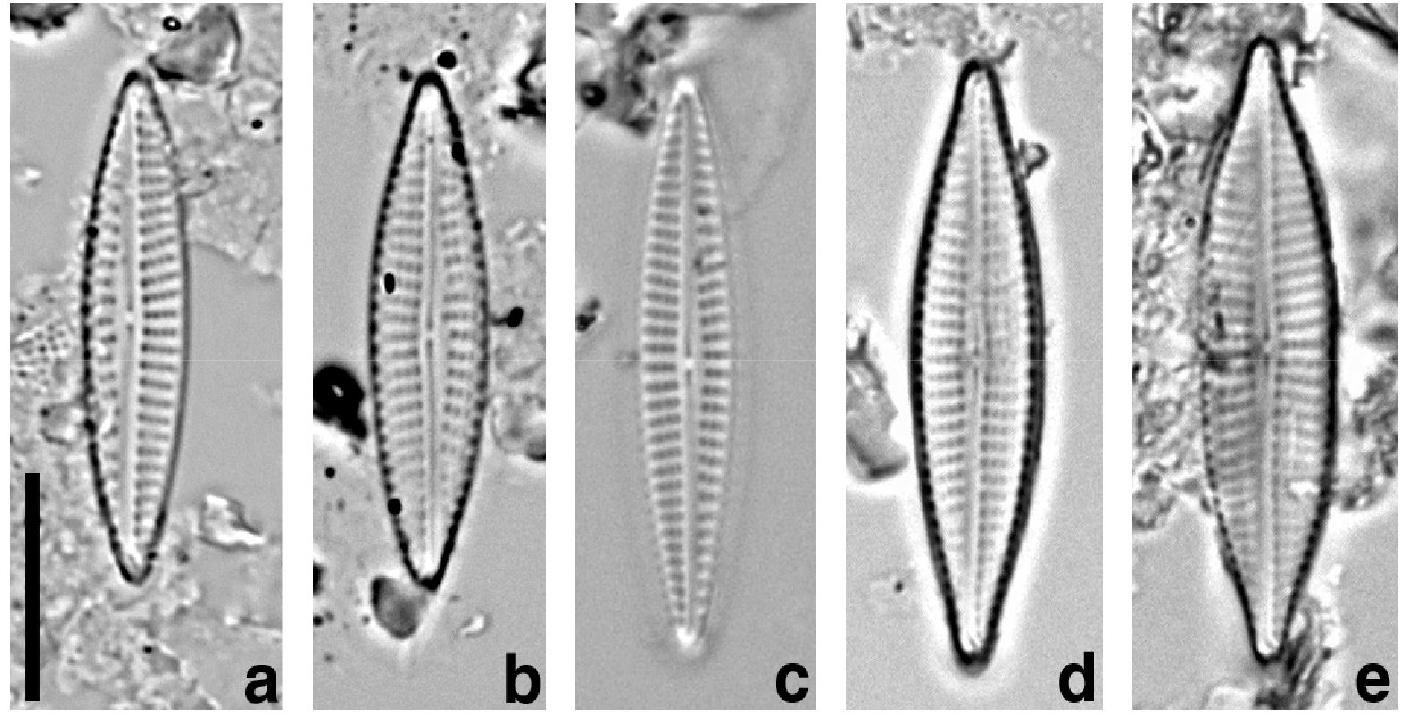
Fig. 1.
LM photos of the Navicula spartinetensis from Yellow sea. (a-b) CH-DD. (c) CH-DL. (d-e) KO-BO. scale bar, 10 ㎛.
주사전자현미경 관찰 : 개각면(valve face)은 거의 평편하다(Fig. 2(a)). 각투(valve mantle)는 얕고, 그물눈은 각투까지 이어진다(Figs. 2(a) and 2(c)). 등줄 가슴판은 두꺼우며, 중앙영역의 등줄 끝에서 개각면보다 살짝 돌출되어 있다(Figs. 2(b-c)). 중앙 영역에서의 등줄 틈(raphe fissure)은 돌출된 부분에서 틈이 넓게 나타난다. 등줄은 직선형이고, 등줄 가슴판은 등줄이 끝나는 부분에서 가로방향으로 약하게 확장된다(Figs. 2(a) and 2(c)). 개각면 양 끝에서 등줄들의 끝(terminal raphe endings)은 같은 방향으로 굽어있다(Figs. 2(a) and 2(c)). 중앙 영역의 그물눈 가로 길이가 양 끝 쪽의 그물눈 가로 길이보다 짧다(Fig. 2(c)).

Fig. 2.
Scanning electron microscopic views of Navicula spartinetensis from Yellow sea. (a), (c) External valve face with elongate areolae. Slightly elevated raphe sternum, terminal raphe endings hooked to the same direction. (b) Central raphe endings simple. (d) Terminal raphe ending.
3.1.2 Navicula flatanica와의 비교
위의 관찰내용을 바탕으로 우리 연구의 출현 종과 N. flanatica의 원 기재 내용을 비교하였다.
N. flanatica (Grunow, 1860)의 원전 기재 내용 : 규각은 좁고 피침형으로, 양 끝은 다소 길쭉하다. 등줄은 직선형으로 중앙의 결절(Central nodule)은 직사각형이다. 규각의 길이는 45 ㎛, 너비는 6.8 ㎛이다. 10 ㎛의 점무늬열 밀도는 11-13개이고, 가로방향 10 ㎛당 그물눈의 밀도는 35개 이상이다. 점무늬열은 방사형으로, 광학현미경에서 그물눈을 뚜렷하게 볼 수 없다.
본 연구에서 출현한 Navicula spartinetensis의 형태적 특징과 N. flanatica의 원전에 기재된 형태적 특징을 비교하였을 때, 가장 뚜렷한 형태적 차이는 점무늬열 밀도이다. 점무늬열의 밀도는 같은 속의 여러 유사한 종들을 나누는 중요한 형태적 특징 중 하나이다. N. flanatica는 점무늬열의 밀도가 10 ㎛에 11-13개인데 비해, N. spartinetensis의 점무늬열 밀도는 10 ㎛당 13-15개를 보였다. 또한 N. spartinetensis 개각의 길이는 19.3-29.3 ㎛로, 45 ㎛인 N. flanatica보다 뚜렷하게 더 작은 개체 크기를 가졌다. 점무늬열 배열에서도 두 종의 차이는 분명하였다. N. spartinetensis의 경우 점무늬열 배열은 중앙영역에서 방사형이며 양 끝 쪽에서는 수렴하는 형태라면, N. flanatica는 전 개각면에 걸쳐서 약한 방사형인 것으로 관찰되었다. 마지막으로 관찰된 뚜렷한 차이점으로서 N. spartinetensis의 경우 광학현미경에서 그물눈의 형태를 뚜렷하게 볼 수 있다.
3.2 동북아시아에서 Navicula spartinetensis의 생물 지리학
N. spartinetensis의 세계적인 분포는 여러 문헌을 통해서 남아메리카와 유럽의 갯벌과 하구 퇴적물에서 서식하는 것을 확인하였다(Méléder et al., 2007; Witkowski et al., 2012). N. spartinetensis의 분포는 앞서 알려진 것보다 더 많은 곳에서 분포할 것으로 예상되나, 명확하게 동정되어 보고된 경우가 많지 않았다. 이는 Navicula 속의 종들 중에서 뚜렷한 특징이 없는 종의 경우, 속 수준까지 동정되는 경우가 비일비재하기 때문이다. 더불어 여전히 많은 지역에서의 규조류 식물상 연구가 부족하다는 것을 의미하기도 한다. 본 연구에서는 N. spartinetensis의 한국에서 분포뿐만 아니라, 더 나아가 동북아시아에서의 분포를 검토하기 위해서 한국, 중국, 그리고 일본에서 기존에 연구한 규조류 문헌을 조사하였다. N. spartinetensis는 이전 연구에서 한국, 중국과 일본에서 보고된 적이 없었으며, 따라서 본 연구를 통해 한국뿐만 아니라 동북아시아에도 최초로 보고되는 바임을 확인하였다. 그리고 추가적으로 N. spartinetensis가 오동정되었을 확률이 높은 N. flanatica의 동북아시아 출현 기록도 함께 검토하였다.
한국의 경우 인천 연안(Cho and Kim, 1988), 금강(김과 조, 1985), 그리고 낙동강 하구(조, 1989, pl. 12, fig. 4)에서 N. flanatica의 출현 기록이 있었다. 또한 새만금(Park, 2011, pl. 12, fig. 14)에서 N. cf. flanatica의 출현 기록이 있었다. 낙동강 하구와 새만금에서 보고된 종의 기재와 도판을 살펴 본 바로는 두 기록 모두 N. spartinetensis의 오동정이었다(Table 2). 인천 연안과 금강에서의 출현 기록에 있어서는 도판 및 기재가 없었으므로 해당 종의 정확한 실체를 확신할 수 없다. 다만 두 기록의 공동 저자였던 조가 낙동강 하구에서 N. spartinetensis를 N. flanatica로 동정한 바 있었으므로 해당 인천 연안 및 금강의 경우에도 N. spartinetensis의 오동정이었을 것으로 여겨진다.
Table 2.
Previous reports of the “Navicula flanatica” from the Northeast Asia with their suggested re-identification
|
Reported
identification
|
Size
(µm)
|
Structures
in 10 µm
|
Striae at
central area
|
Striae at
apices area
|
Areolae
under LM
|
Distribution
|
Suggested
re-identification
|
| N. flanaticaa |
L:27.3
W:5.3
|
S:12
A:20
|
Radiate
|
Convergent
|
Conspicuous
|
South Sea, Korea
| N. spartinetensis |
| N. flanaticab |
L:24.5
W:6
|
S:11
A:20
|
Radiate
|
Convergent
|
Conspicuous
|
Yellow Sea, Korea
| N. spartinetensis |
| N. flanaticac |
L:41-61
W:9-12
|
S: 9-10
A:>20
|
Slightly radiate
|
Radiate
|
Inconspicuous
|
Ariake Sea, Japan
| N. flanatica |
| N. flatanicad |
L:26.5
W:6
|
S:12
A:20
|
Radiate
|
Convergent
|
Conspicuous
|
Ariake Sea, Japan
| N. spartinetensis |
일본에서는 아리아케해(有明海) 나나우라(七浦) 갯벌(Park et al., 2012, fig 7-L)과 이사하야(諫早) 갯벌(Ohtsuka, 2005, fig. 5)에서 N. flantanica가 보고된 바 있다(Table 2). 해당 종명은 적법하지 않은 명칭으로서, N. flanatica의 오타이다. 나나우라 갯벌에서 보고된 N. flanatica의 경우 원전에 기재된 바와 일치하는 형태적 특징을 보였다. 한편 이사하야 갯벌에서 보고된 ‘N. flanatica’의 경우 N. spartinetensis과 잘 일치하는 형태적 특징을 보이고 있다. 따라서 N. spartinetensis는 동북아시아 특히 황해 갯벌 연안에 널리 분포하고 있는 것으로 여겨진다(Fig. 3).
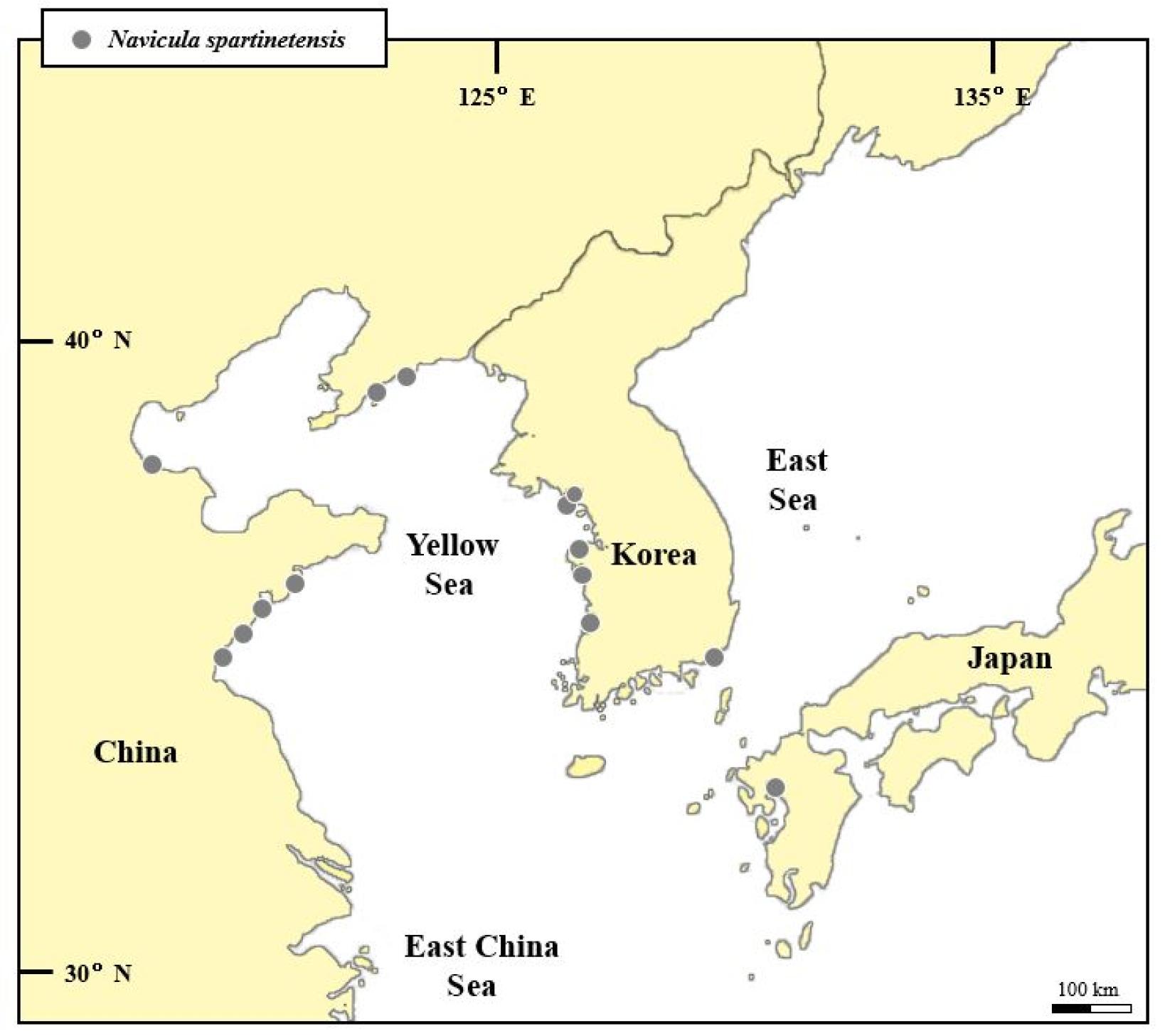
Fig. 3.
Biogeographic distribution of Navicula spartinetensis in Northeast Asia.
4. 결 론
국내 규조류 분류 연구에 있어 2000년 대 이전까지는 주로 광학현미경 관찰을 통해서 종을 동정하였다. 광학 현미경을 통한 동정 과정에서 국내보다 규조류 연구사가 긴 유럽과 미국 등의 해외 문헌에 의존하였으며, 국내 규조류 군집 고유의 다양성 보다는 유럽 및 미국의 규조류 군집과의 유사성이 부각되는 연구가 이루어질 수 밖에 없었다. 2000년 대 이후로는 광학현미경뿐만 아니라 미세구조 관찰에 용이한 전자현미경 사용이 일반화되었으며 그에 따라 국내 규조류 미기록종에 대한 연구가 활발해졌다. 그러나 기존에 보고된 종들에 대한 분류학적 재검토는 거의 이루어지지 않고 있다. 본 연구에서는 국내에서 흔히 N. flanatica로 동정되었던 종에 대한 분류학적 재검토를 수행하였다. 광학현미경 및 전자현미경 관찰 결과 해당 종의 정확한 동정이 N. spartinetensis임을 밝혔는데, 해당 종의 출현 기록은 국내뿐 아니라 동북아시아에서도 최초이다. 이번 연구를 통해서 유럽과 미국의 연구 결과에 의지하였던 과거의 규조류 동정에 오류가 있음이 확인되었으며, 향후 분류학적 재검토가 반드시 이루어져야 할 것이다.
Acknowledgements
본 연구는 한국연구재단 신진연구지원사업(NRF-2018R1C1B6002832) 지원을 받아 수행되었음.
References
김준호, 조경제, 1985. 금강하구 조간대 저토의 물리화학적 특성, 저생규조의 종조성 및 생물량. 한국생태학회지, 8: 21-29.
김학균, 1989. 마산만의 와편모적조의 발생과 환경특성. 국립수산진흥원 연구보고, 43: 1-40.
10.1515/hfsg.1989.43.1.40박주석, 1979. 마산만의 환경오염 평가를 위한 야외생물 오염 시험연구. 한국해양학회지, 14: 15-25.
박주석, 1980. 한국 남해안의 식물성 플랑크톤의 출현양 및 조성과 이들이 먹이와 적조로써 양식생물에 미치는 영향. 국립수산진흥원 연구보고, 23: 7-157.
박주석, 1982. 진해만 적조의 특성과 환경변화. 국립수산진흥원 연구보고, 28: 55-58.
10.1159/000212592박태수, 1956a. 한국해협에 있어서 플랑크톤의 계절적 변화에 관하여 (I). 부산수산대학 연구보고집, 1: 1-12.
박태수, 1956b. 하기 남해안에 있어서 Microplankton의 양 및 조성에 관한 연구. 부산수산대학 연구보고집, 1: 13-32.
유광일, 이종화, 1976. 마산만의 환경학적 연구. 2. 식물성플랑크톤의 년변화. 한국 해양학회지, 11: 34-38.
유광일, 1973. 백령도의 식물성 플랑크톤. 한양대학교 문리대 학보, 28: 461-462.
유광일, 1974. 식물성 플랑크톤의 종조성과 현존량. - 다도해의 굴성장요인과 연해수질 오염에 관한 연구 -. 대한민국 학술원 논문집(자연과학편), 13: 326-329.
유광일, 1980. 오염해역에 있어서의 생물군집의 환경지표성에 관한 연구. 한양대학교 환경과학연구소 논문집, 1: 10-14.
윤양호, 노경희, 김영기, 1991. 제주도 연안해역에 출현하고 있는 적조생물. 제주대학교 해양연구소 연구보고, 15: 1-14.
이민재, 심재형, 김종균, 1967. 한국 해역의 플랑크톤 연구. 하기 서해에 있어서의 해황과 식물성 플랑크톤 관하여. 서울대학교 해양생물연구소, 1: 1-14.
이진환, 조경제, 2015. 국가 생물종 목록집. 국립생물자원관, 인천, 365 pp.
정영호, 심재형, 이민재, 1971. 하계 경기만의 수질오염과 생산력에 관한 연구. 제3보. 식물성 플랑크톤의 분류 (II). 한국식물학회지, 14: 47-59.
조경제, 1989. 낙동강 하구 저토의 환경구배에 따른 저서규조류의 군집구조. 박사학위논문. 서울대학교, 서울, 106 pp.
최 상, 1967. 한국 해역의 식물플랑크톤에 관한 연구. II. 한국 연안수역의 식물플랑크톤. 한국해양학회지, 2: 1-12.
최 상, 1972. 고리 해역의 부유생물과 유기현탁물질의 소장. 한국해양학회지, 7: 47-58.
최정신, 1969. 수영만에 있어서 규조류의 계절적인 양 및 조성의 변화. 한국수산학회지, 2: 16-24.
Bourasi, S.K., S.D. Singh, P. Patil and P. Rathor, 2018. A study of effect of Physico-chemical parameters on biodiversity of Diatoms special Reference to
Navicula in river Narmada at Harda, MP India. Indian Journal of Life Sciences, 6(2): 575-578.
Cho, C.H., 1981. On the
Gymnodinium red tide in Jinhae Bay. Bulletin of the Korean Fisheries Society, 14: 227-232.
Cho, K.J. and J.H. Kim, 1988. Species composition and Primary production of the Benthic algal assemblage along a channel in salt marsh, Kyonggi bay, Korea. The Korean Journal of Ecology, 11(1) : 1-15.
Choi, J.H., 1991. Estimation of Boundary Shear Velocities from Tidal Current in the Gyeonggi Bay, Korea. Journal of the Korean Society of Oceanography, 26: 340-349.
Gligora M., K. Kralj, A. Plenković-Moraj, F. Hinz, E. Acs, I. Grigorszky, C. Cocquyt and B. Van de Vijver, 2009. Observations on the diatom
Navicula hedinii Hustedt (Bacillariophyceae) and its transfer to a new genus
Envekadea Van de Vijver et al. gen. nov. European Journal of Phycology, 44(1): 123-138.
10.1080/09670260802389783Grunow, A., 1860. Über neue oder ungenügend gekannte Algen. Erste Folge, Diatomeen, Familie Naviculaceen. Verhandlungen der Kaiserlich-Königlichen Zoologisch-Botanischen Gesellschaft in Wien, 10: 503-582.
Guiry, M.D. and K. Gandhi, 2019. Decussiphycus gen. nov.: a validation of "Decussata" (R.M. Patrick) Lange-Bertalot (Mastogloiaceae Bacillariophyta). Notulae Algarum, 94: 1-2.
Koh C.-H. and J.S. Khim, 2014. The Korean tidal flat of the Yellow Sea: Physical setting, ecosystem and management. Ocean & Coastal Management, 102; 398-414.
10.1016/j.ocecoaman.2014.07.008Kokubo, S., 1940. Quantitative reports of plankton in Japanese coasts. Plankton Jibo, 15: 121-154 (in Japanese).
Licea S, J.L. Moreno-Ruiz and R. Luna, 2016. Checklist of Diatoms (Bacillariophyceae) from the Southern Gulf of Mexico: Data-Base (1979-2010) and New Records. Journal of Biodiversity & Endangered Species, 4(3): 1-7.
Liu Y., J.P. Kociolek, Y. Fan and Q. Wang, 2012.
Pseudofallacia gen. nov., a new freshwater diatom (Bacillariophyceae) genus based on
Navicula occulta Krasske. Phycologia, 51: 620-626.
10.2216/11-098.1Méléder, V., Y. Rincé, L. Barillé, P. Gaudin and P. Rosa, 2007. Spatiotemporal changes in microphytobenthos assemblages in a macrotidal flat (Bourgneuf Bay, France). Journal of Phycology, 43: 1177-1190.
10.1111/j.1529-8817.2007.00423.xMitbavkar, S. and A.C. Anil, 2002. Diatoms of the microphytobenthic community: population structure in a tropical intertidal sand flat. Marine Biology, 140: 41-57.
10.1007/s002270100686Oh, J.K. and B.C. Kum, 2001. Depositional of marine environmental engineering, 9, 1-13.l Environments and Characteristics of Surface Sediments in the Nearshore and Offshore off the Mid-Western Coast of the Korean Peninsula. Journal of the Korean earth science society, 22: 377-387.
Oh, S.H. and C.H. Koh, 1995. Distribution of diatoms in the surficial sediments of the ManKyung Dongjin tidal flat, west coast of Korea (Eastern Yellow Sea). Marine Biology, 122: 487-496.
10.1007/BF00350883Ohtsuka, T., 2005. Epipelic diatoms blooming in Isahaya Tidal Flat in the Ariake Sea, Japan, before the drainage following the Isahaya-Bay Reclamation Project. Phycological Research, 53: 138-148.
10.1111/j.1440-1835.2005.tb00365.xPark, J., B.-O. Kwon, M. Kim, S. Hong, J. Ryu, S.J. Song and J.S. Khim, 2014. Microphytobenthos of Korean tidal flats: A review and analysis on floral distribution and tidal dynamics. Ocean & Coastal Management, 102: 471-482.
10.1016/j.ocecoaman.2014.07.007Park, J., 2011. Flora of Marine Benthic Diatoms from the Saemangeum Tidal Flat of Korea. Ph.D.Thesis, Seoul National University, Seoul, 195 pp.
Park, J. and C.-H. Koh, 2012. Taxonomic studies on Korean marine benthic diatoms LM and SEM observations of the diatom genus Amphora from Korean tidal flats. Ocean Science Journal, 47: 101-112.
10.1007/s12601-012-0011-9Park, J., J.H. Lee and J.S. Khim, 2016. The Identity of '
Berkeleya scopulorum' from Northeast Asia: Report on
Climaconeis mabikii sp. nov. from Temperate Marine Waters with Notes on Biogeography of the Genus. Ocean Science Journal, 51: 591-598.
10.1007/s12601-016-0059-zPark J.S., J.S. Khim, T. Ohtsuka, H. Araki, A. Witkowski and C.-H. Koh, 2012. Diatom assemblages on Nanaura mudflat, Ariake sea, Japan: with reference to the biogeography of marine benthic diatoms in Northeast Asia. Botanical Studies, 53: 105-124.
Round, F.E., R.M., Crawford and D.G. Mann, 1990. The Diatoms. Cambridge, New York. 747 pp.
Skvortzow, B.W., 1929. Freshwater diatoms from Korea, Japan. Philip. Philippine Journal of Science, 38: 283-291.
Skvortzow, B.W., 1932. Diatoms from the bottom of the sea of Japan. Philippine Journal of Science, 47: 265-280.
Sullivan, M.J. and C.W. Reimer, 1975. Some diatoms (Bacillariophyceae) from a Delaware salt marsh - four of which are described as new. Botanica Marina, 18: 115-121.
10.1515/botm.1975.18.2.115Underwood, G.J.C. and J. Kromkamp, 1999. Primary production by phytoplankton and microphytobenthos in estuaries. Advances in ecological research, 29: 93-153.
10.1016/S0065-2504(08)60192-0Witkowski, A., M.J. Sullivan, B. Bogaczewicz-Adamczak, M. Bąk, E. Rhiel, L. Ribeiro and P. Richard, 2012. Morphology and distribution of a little known but widespread diatom (Bacillariophyceae), Navicula spartinetensis Sullivan et Reimer. Diatom Research, 27(1): 43-51.
10.1080/0269249X.2011.647772Yoo, K.I. and J.H. Lee, 1979. Environmental studies of the Jinhae Bay. 1. Annual cycle of phytoplankton population, 1976-1978. Journal of Oceanological Society of Korea, 14: 26-31.
Yool, A. and T. Tyrrell, 2003. Role of diatoms in regulating the ocean's silicon cycle. Global Biogeochemical Cycles, 17: 1103.
10.1029/2002GB002018

