

1. 서 론
갯벌은 서해안과 남해안에서 우리나라 연안 생태계를 특징짓는 대표적이고도 중요한 생태계의 하나이다. 최근 들어 국내외 다양한 연구자들에 의해 습지에 대한 연구가 진행됨에 따라, 연안습지인 갯벌이 보유하고 있는 중요한 생태계에서의 기능이 점차 밝혀지고 있다(Yu et al., 2010). 갯벌이 포함되어있는 연안지역은 다양한 해양생물들의 서식 및 산란 등이 집중적으로 이루어지는 생태적으로 매우 중요한 서식처이며(Potter et al., 1983; Van der Veer et al., 1990; Santos and Nash, 1995; Nash and Santos 1998; Fujita et al., 2002), 높은 일차생산력으로 인하여 서식하고 있는 생물들이 이용 가능한 먹이원이 풍부하다는 특징이 있다는 것도 밝혀졌다(Levinton, 1995). 더불어 우리나라 서해안에 넓게 형성되어 있는 갯벌은 다양한 지리적, 퇴적학적, 해양 환경적 특징을 가지고 있으며, 그에 따라 이에 적합한 다양한 생물들의 서식처 역할을 하고 있다(Woo et al., 2005).
저서 환경에 직․간접적 영향을 받는 생물들 중 중형저서동물(meiofauna)은 대체로 망목크기 1 mm의 체를 통과하여 32 µm의 체에 걸러지는 저서생물이다(Shirayama, 1993). 중형저서동물에 속하는 동물군의 주요 종들은 전 생활사를 퇴적물 안에서 보내며 서식을 위해 비교적 안정된 환경을 요구하고, 세대기간이 짧기 때문에, 대형저서동물에 비하여 환경 변화에 민감하여 짧은 기간에도 군집의 변화를 보인다고 알려져 있다(Sandulli and De Nicola, 1990). 단위 면적당 서식밀도가 퇴적물 1 m2 당 106∼107으로 다른 생물 그룹들에 비해 상대적으로 아주 높아 밀도 변화의 감지가 용이하고(Mcintyre, 1969), 조사 방법에 있어서도 매우 적은 시료채취만으로도 분석이 가능하기 때문에, 시료 채집으로 인한 자연환경의 훼손이 적다는 특징이 있다(Moore and Bett, 1989). 우리나라에서도 해양 생태계 내에서의 중형저서동물 군집의 중요성에 대한 인식이 점차 증대되고 있지만, 아직은 소수의 연구자들에 의해 국내 해양에서의 중형저서동물에 대한 연구들이 이루어지고 있다(Kim et al., 1998; Kim et al., 2000; Kim et al., 2001; Kim et al., 2004a; Kim et al., 2004b; Min et al., 2006; Back et al., 2009; Kang et al., 2011; Kim et al., 2014).
본 연구에서는 서해안 근소만의 중형저서동물 군집에 관한 기초자료를 모으고, 근소만의 서식처로서의 생태학적 가치를 이해하기 위하여 중형저서동물의 계절별 군집 변동 양상을 파악하고자 하였다. 본 연구의 대상 지역인 충청남도 태안반도에 위치한 근소만 갯벌은 조석의 영향을 받는 반폐쇄성 만으로 총면적은 30 ㎢이며, 이 중 70%가 저조시 대기 중에 노출된다(Kim and Kim, 2008; Choi et al., 2010). 조간대 퇴적물은 주로 사니질로 구성되어 있고 조차는 6 m이며, 만조 시에는 수심이 2~4 m 정도이다(Kim and Kim, 2009). 근소만은 직접 유입되는 하천이 없기 때문에 만 외부와 내부의 해수교환은 약 2 ㎞ 폭의 만 입구를 통해서만 이루어진다(Kim and Kim, 2008). 이 연구에서는 근소만에서의 현장조사를 통해 퇴적물을 채집하고 실험실에서 중형저서동물의 군집 특성을 분석하여, 약 1년간에 걸쳐 나타난 연구지역 내 중형저서동물 군집의 계절별 변화 양상을 파악하고 군집구조의 특성을 분석하는데 중점을 두었다. 또한 연구지역 내 각 정점별․계절별 저서생태계의 건강도를 평가하고자 선충류(Nematodes)와 저서성 요각류(Harpacticoids)의 서식밀도 비를 이용한 생물지수인 N/C ratio와 중형저서동물 분류군 중에서 매우 우점하는 선충류의 섭식 유형을 이용하여 오염도를 평가하는 ITD Index를 활용하였다.
재료 및 방법
태안 근소만에서의 중형저서동물 군집 분석을 위하여 근소만 내 조간대 중부에 St. 1과 2를 설정하였으며, 상부에 St. 3과 4를 설정하였다. 퇴적물 시료 채집은 2013년 8월, 10월, 2014년 1월, 4월에 이루어졌다(Fig. 1). 퇴적물 시료 채집 정점은 근소만 내의 조간대 중부에서 St. 1과 2를 채집하였고, 상부에서 St. 3과 4를 채집하였다. 퇴적물 시료는 직경 3.6 cm의 아크릴코어러를 이용하여 각 정점에서 반복수 3으로 채집하였다. 채집한 시료는 퇴적물 표층으로부터 3 cm까지를 1 cm간격으로 자른 후, 로즈 벵갈(rose bengal)을 혼합한 5% 중성 포르말린으로 고정하여 연구실로 운반하였다. 중형저서동물의 분포는 일반적으로 퇴적물 상층 3 cm 내에 90% 이상을 차지하고 있어(Azra Ansari, 2001; Kang et al., 2011), 퇴적물 상층 3 cm까지의 시료를 분석하였다. 운반된 퇴적물 시료는 실험실에서 1mm와 38 µm 체로 체질하여, 루독스(Silica-gel Ludox HS-40)를 이용한 추출법을 사용하여 퇴적물에서 생물만을 분리하였다(Burgess, 2001). 분리된 중형저서동물은 각 크기의 체(500 µm, 250 µm, 125 µm, 68 µm, 38 µm)를 이용해 크기별로 걸러 광학현미경 하에서 계수 후 분석하였다. 중형저서동물의 생체량은 Shirayama(1983)의 주요 분류군별 개체당의 Ash free dry weight (µg) 환산값을 이용하여 나타내었다.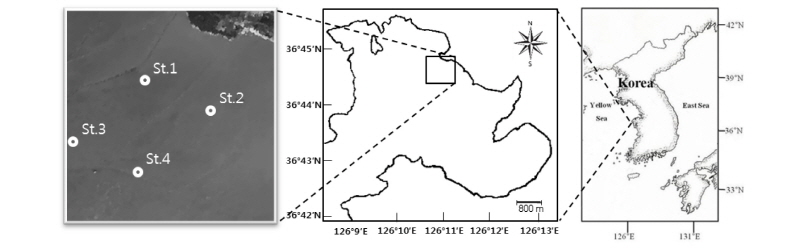
입도분석은 채집한 시료 중 5 g을 비커에 담아 10%의 H2O2로 유기물을, 0.1 N-HCL로 탄산염을 제거한 후, 시료를 4 ø 체로 조립질과 세립질로 구분하여 4 ø 이하의 사질 시료는 Ro-tap sieve shaker로 15분간 체질한 후, 입도 등급별로 무게 백분율을 구하였고, 4 ø 이상의 니질 시료 2 g에 0.1% calgon 용액을 넣고 교반 시킨 후 X-선 자동입도 분석기인 Sedigraph 5000D를 이용하여 입도 무게 백분율을, Folk and Ward(1957)의 Inclusive Graphic Method에 의하여 변수를 구하였다.
채집 시기별 군집구조와 유사도 등을 파악하기 위하여 Bray-Curtis의 유사도 지수를 기초로 집괴분석 및 SIMPROF 분석을 실시하여 군집 특성을 확인하였다(Primer v.6). 각 시료의 계절별․정점별에 따른 중형저서동물 서식밀도의 유의한 차이를 파악하기 위하여 two-way ANOVA test를 실시하였다(SPSS. v.19).
N/C ratio (Nematodes/Harpactidoids)
Raffaelli and Mason, 1981.
N/C (Nematode/Harpacticoids) ratio는 중형저서동물 그룹 중 가장 우점하는 선충류와 저서성 요각류의 서식밀도비를 이용한 지수이다. 선충류는 오염 특히 빈산소 환경에 대한 내성이 강하고 저서성 요각류는 오염에 민감한 생태적 특징을 이용한 평가이다. 값이 높을수록 오염도가 증가하고, 값이 낮을수록 오염도의 감소를 지시한다. N/C ratio 값 산출은 아래와 같은 공식을 이용하여 산출하였다.
N/C ratio = Abundance of Nematodes /Abundance of Harpacticoids
ITD (Index of Trophic Diversity) 지수
Heip et al., 1985.
ITD (Index of Trophic Diversity) 지수는 중형저서동물 그룹 중 가장 우점하는 선충류의 섭식유형을 이용한 지수이다. 선충류를 4가지 식성으로 구분하고(Preben Jensen 1987), 값이 0.25∼1.0의 범위 값을 갖는다. ITD 지수는 값이 높으면 오염도가 증가하고, 값이 낮으면 오염도의 감소를 지시한다. ITD 지수 값의 산출은 아래와 같은 공식을 이용하여 산출하였다.
ITD = Σθ²
(θ = 각 섭식유형의 선충류(Nematodes)서식밀도/선충류(Nematodes)서식밀도)
·no buccal cavity or a fine tubular one - Selective deposit (bacterial) feeders (1A)
·large but unarmed buccal cavity - Non-selective deposit feeders (1B)
·buccal cavity with scraping tooth of teeth - Epistrate (diatom) feeders (2A)
·buccal cavity with large jaws - Predators/omnivores (2B)
결과 및 고찰
서식환경
각 정점에서의 퇴적물 입자의 구성 비율(Compisotion), 평균 입도(Mean Size), 분급도(Sorting), 왜도(Skewness), 첨도(Kurtosis)는 Table 1에 나타내었다. 근소만 지역 내 표층 퇴적물 분석은 St. 1, 2와 St. 3, 4가 같은 정선에 위치하기 때문에 계절별 각 정선을 측정하여 퇴적물 분석을 실시하였다. 중형저서동물의 서식밀도에 영향을 미치는 환경요인에는 온도, 입도, 퇴적물 내 유기물의 양, 저질 간극의 공간, 간극수 내의 산소량 등이 있고(Sandulla and De Nicola, 1990; James and Mark, 2004), 그중 퇴적물의 입도는 저서동물의 분포에 영향을 미치는 중요한 요인이다(Warwick and Buchanan, 1970).
연구지역의 계절별 퇴적물 조성비를 보면, 대부분의 정점에서 Sand의 비율이 Silt보다 높게 나타났으며, 특히 하계(2013년 7월), 동계(2014년 1월), 춘계(2014년 4월)의 St. 1, 2의 Sand 비율은 70% 이상으로 구성되어졌다. 그러나 동계(2014년 1월), 춘계(2014년 4월)의 St. 3, 4의 퇴적물 조성비를 보면, Silt의 비율이 Sand보다 높게 나타났다. St. 3, 4에서의 Slit 비율은 여름부터 점점 높아지는 경향을 나타냈고, St. 1, 2에서는 Sand의 비율이 증가하는 경향을 보였다. 각 정점에서의 평균 입도는 St. 1, 2에서 4.58∼6.35 ø, St. 3, 4에서 3.65∼4.52 ø 값의 범위를 나타냈다. 분급도는 2014년 10월에 St. 1, 2에서 2.08ø 의 값으로 ‘Very poorly sorted' 단계를 나타내었고 그 외 계절에서는 1.52∼1.99ø로 ‘Poorly sorted' 단계를 나타냈다. 본 연구 지역의 평균 입도는 전체적으로 서해에서 Kim et al. (2004)이 조사한 대부도 조간대 지역의 평균 입도인 3.16∼3.73 ø 보다 세립한 특성을 보이고, 조하대 지역인 새만금 해역의 평균 입도 1.7∼5.8 ø (Kim et al., 2014)와 장봉도 해역 평균입도 1.49∼3.55ø (Kang et al., 2011) 보다 세립한 형태를 보이고 있다.
중형저서동물 서식밀도
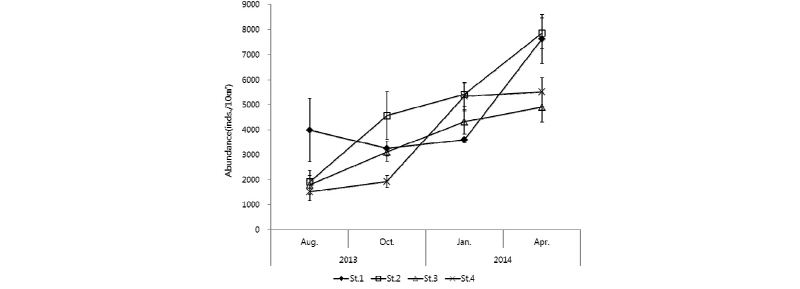
각 조사 시기의 각 정점에서 중형저서동물의 서식밀도를 살펴보면, 2014년 4월에 모든 정점에서 가장 높게 나타났고, 2013년 8월에 St. 1을 제외한 모든 정점에서 가장 낮게 나타났다(Fig. 2). 중형저서동물 평균 서식밀도는 2014년 4월에 St. 2에서 7,849 inds./10 cm2로 가장 높은 값을 보였고, 2013년 8월 St. 4에서 1,521 inds./10 cm2으로 가장 낮은 값을 보였다(Table 2). 중형저서동물의 군집 분포에 영향을 주는 주요 요인 중 하나로 계절적 변동을 들 수 있는데, 중형저서동물의 일반적인 계절적 분포 양상은 여름철에 높은 서식밀도를 보이고 겨울철에는 낮은 온도 등의 영향으로 서식밀도가 감소하는 경향을 나타낸다고 알려져 있다(James and Mark, 2004). 각 정점별로 중형저서동물 서식밀도의 계절적 변동을 보면 St. 2, 3, 4에서는 하계에서 동계를 거쳐 춘계로 갈수록 서식밀도가 증가하였고, St. 1에서의 중형저서동물 서식밀도는 추계와 동계에 감소하다가 동계에서 춘계로 계절이 변할 때, 서식밀도 값이 증가하는 양상을 나타냈다. 중형저서동물의 서식밀도에 있어서 계절적 변동은 모래 퇴적물보다는 펄로 구성된 퇴적물에서 더욱 변화가 크게 나타난다(Bruce, 1985).
각 정점별 중형저서동물 서식밀도를 보면(Fig. 2, Table 2), St. 1에서의 중형저서동물 서식밀도는 2013년 8월보다 10월에 다소 낮아지면서 가장 낮은 값을 보였고, 2014년 1월에는 소폭으로 증가하였고 4월에 급격하게 증가하는 양상을 보였다. St. 2에서의 중형저서동물 서식밀도는 2013년 10월에 크게 증가하였고, 그 이후 2014년 1월에 소폭으로 증가하였다가 다시 4월에 크게 증가하였다. St. 3에서의 중형저서동물 서식밀도는 계절이 지나면서 서서히 증가하였으며, St. 4에서는 2013년 8월보다 10월에 서식밀도가 약간 증가하였고 2014년 1월과 4월에는 급격하게 증가하는 양상을 나타내었다. 중형저서동물 서식밀도는 St. 1을 제외한 St. 2, 3, 4에서 여름에 가장 낮은 값을 보이고 계절이 변할수록 서식밀도가 증가하는 값을 나타냈다. 일반적으로 온도가 여름에 높아지고 겨울에 낮아지는 환경변화에 동반하여, 중형저서동물 서식밀도 값도 여름에 높고 겨울에 낮은 것이 서해안 갯벌에서의 일반적인 중형저서동물 서식밀도의 계절적 변동 현상이다(Kim et al., 2005; Min et al., 2006; Kang et al., 2011). 본 연구에서 일반적인 중형저서동물 서식밀도의 계절변동 양상과 다른 결과가 나온 것에 대해서는 여러 요인이 있을 수 있겠지만, 연구지역인 근소만은 바지락이 넓게 분포되어 서식하고 있어 채집을 위하여 사람들이 많은 왕래를 하고 있다는 것과, 특히 여름에 시민단체나 일반인들에 의한 갯벌 체험에 의해서 훼손 등에 의한 인위적인 영향을 생각해 볼 수 있을 것이다(Kim and Kim, 2010).
각 계절별 중형저서동물 서식밀도를 정점 간에 비교해보면(Fig. 2, Table 2), 2013년 8월 하계 조사 시에는 St. 1에서 3,984 inds./10 cm2로 가장 높았고, St. 4에서 1,521 inds./10 cm2로 가장 낮았다. 2013년 10월 추계 조사 시에는 St. 2에서 4,565 inds./10 cm2로 가장 높았고, St. 4에서 1,929 inds./10 cm2로 가장 낮았다. 2014년 1월 동계 조사 시에는 St. 2에서 5,407 inds./10 cm2로 가장 높았고, St. 1에서 3,594 inds./10 cm2로 가장 낮았다. 2014년 4월 춘계 조사 시에는 St. 2에서 7,849 inds./10 cm2로 가장 높았고, St. 3에서 4,902 inds./10 cm2로 가장 낮은 서식밀도 값을 나타냈다.
전체 중형저서동물 서식밀도는 정점별‧계절별로 유의한 차이를 보였다(Two way ANOVA, p<0.05). 본 연구에서 중형저서동물 서식밀도는 모든 정점에서 1,521∼7,849 inds./10 cm2 범위로 평균 4,161 inds./10 cm2의 값을 나타내었다. 서해안 다른 지역에서의 중형저서동물 서식밀도 연구결과를 보면, 대부도 갯벌에서 30∼1,382 inds./10 cm2, 평균 751 inds./10 cm2, 새만금갯벌에서 29∼4,107 inds./10 cm2, 평균 1,020 inds./10 cm2, 이원방조제 갯벌에서 246∼2,177 inds./10 cm2, 평균 870 inds./10 cm2로 보고되어 있다(KORDI, 2001; Kim et al., 1998; Min et al., 2006). 본 연구에서의 중형저서동물 서식밀도는 기존 서해안에서 연구된 중형저서동물 서식밀도 연구 결과보다 높은 서식밀도를 보이고 있다. 이는 이 지역의 중형저서동물이 다른 해역의 환경조건보다 서식하기에 적합한 주변 해양환경을 가지고 있다는 것을 의미한다고 보여진다. 중형저서동물은 퇴적물 내의 박테리아, chlorophyll a와 밀접한 관계가 있다고 알려져 있다(Danovaro et al., 1999; Grove et al., 2006; Kim et al., 2014). Choi et al.(2010)에 의하면 2007년 2월과 5월의 chlorophyll a의 농도 값이 본 연구지역의 St. 1, 2와 인접한 정점에서는 2월에서 근소만 전 지역의 측정 농도에 비교하였을 때 매우 높은 농도를 보이다가 5월에 매우 낮은 농도로 변하였고, St. 3, 4와 인접한 정점에서는 근소만 전 지역의 측정 농도의 평균값과 유사한 농도이며 2월보다 5월에 소폭 낮아진 농도를 나타내었다. 본 연구 결과에서는 St. 1, 2에서 2014년 1월에서 4월로 가면서 매우 높은 폭으로 서식밀도 값이 높아졌고, St. 3, 4에서는 소폭 증가한 서식밀도 값을 나타내었는데, 이와 비교해봤을 때는 chlorophyll a의 농도와 중형저서동물의 서식밀도 값이 음의 상관관계를 보였지만, 같은 시기 및 정점에서 조사된 것이 아니기 때문에, 향후 동일한 시기와 정점에 chlorophyll a의 농도와 중형저서동물의 서식 밀도 값을 측정 및 분석하여 보다 확실한 연구 결과를 도출할 필요가 있을 것이다.
우점분류군 서식밀도
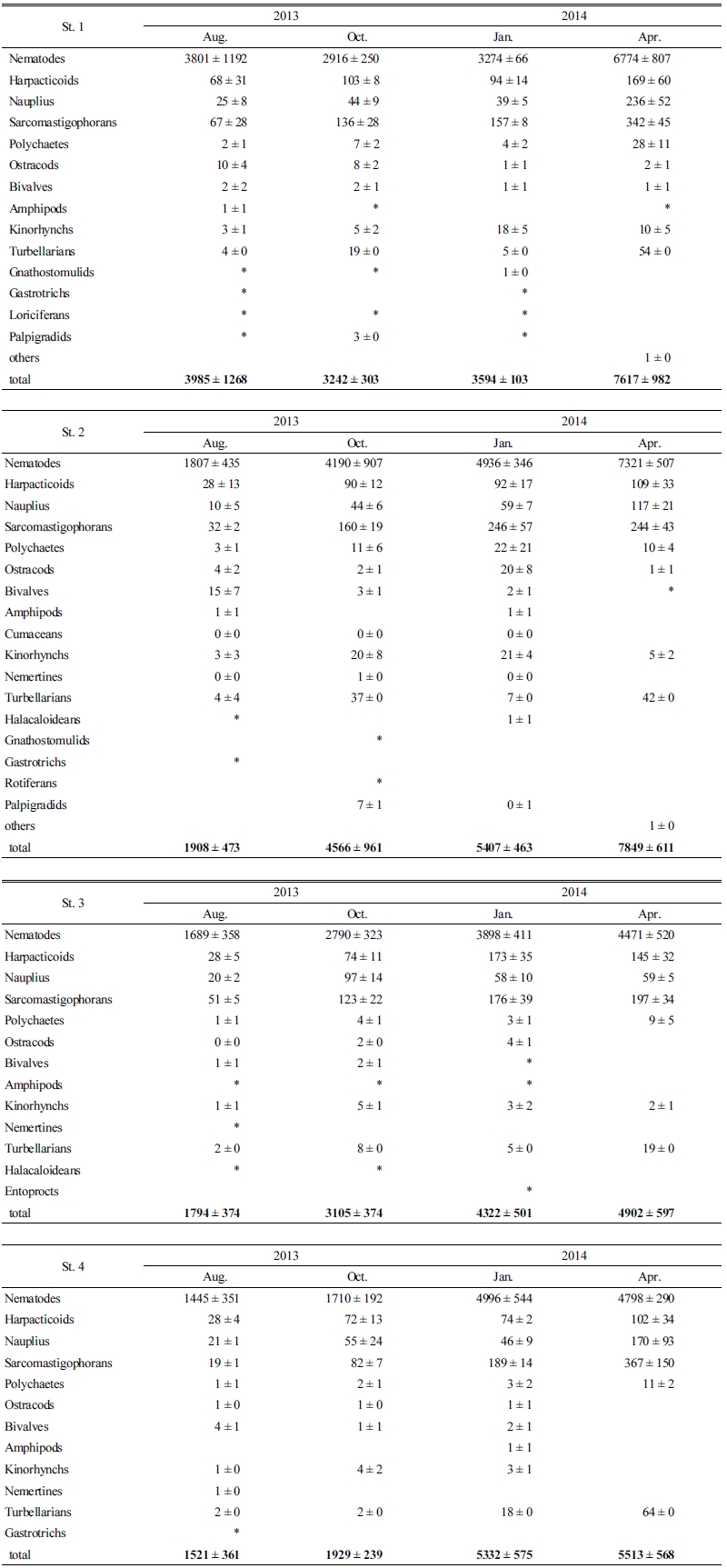
본 연구에서 정점별‧계절별 출현한 중형저서동물 분류군 중 가장 우점하여 나타난 분류군은 선충류(Nematodes)로 나타났으며(Fig. 3, Table 2), 출현 분류군 중 선충류의 비율은 90.6∼92.5%였다. 선충류는 일반적으로 중형저서동물 분류군 중에서 비교적 개체 수가 많고, 종 다양성 또한 높은 분류군으로 알려져 있고(James and Mark, 2004), 일반적으로 조간대의 세립질 퇴적물에서는 선충류가 우점하는 경향이 많은 것으로 알려져 있다(Giere, 1993). 우리나라 서해의 서쪽 해역인 중국 Shandong과 Jiangsu 연안의 대륙붕 해역에서 조사된 중형저서동물 군집 결과에 나타난 선충류 비율이 73.8∼92.8%로 나타난 것과 비교하면 약간 높게 나타났으나, 큰 차이는 보이지 않았다(Xiao-Shou Liu et al., 2007). 선충류 다음으로 우점하여 출현하는 분류군은 정점별 차이는 있지만 저서성 유공충류(Sarcomastigophorans), 저서성 요각류(Harpac-ticoids), 갑각류 유생(Nauplius), 편형동물(Turbellarians) 순이었다. 이들 5개 분류군의 서식밀도 비율은 전체 중형저서동물 서식밀도의 95% 이상을 차지하는 것으로 나타나 일반적인 해양 저서환경에서 나타나는 출현 분류군과 큰 차이를 보이지는 않았다. Dorris et al.(1999)에 따르면, 일반적으로 서식밀도가 높은 중형저서동물 분류군은 선충류, 저서성 요각류, 편형동물, 동문동물, 완보동물, 복모동물이 있고 이들은 대부분의 해역에서 나타나는 것으로 알려져 있다.
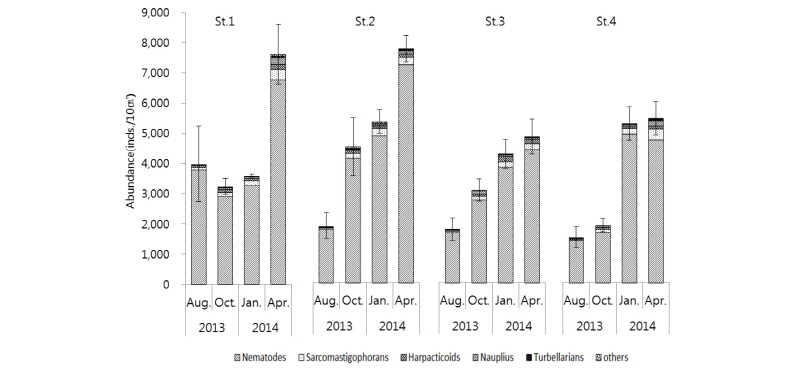
최우점 분류군인 선충류의 계절별 평균 서식밀도는 2014년 4월에 가장 높은 5,841 inds./10 cm2가 출현하였고, 2013년 8월에 가장 낮은 2,186 inds./10 cm2가 출현하였다. 선충류 다음으로 우점하는 저서성 유공충류의 평균 서식밀도는 2014년 4월에 가장 높은 288 inds./10 cm2가 출현하였고, 2013년 8월에 가장 낮은 42 inds./10 cm2가 출현하였다. 저서성 유공충류의 계절별 서식밀도는 St. 3을 제외하고 모든 정점에서 2014년 4월에 가장 높게 나타났고 그 다음으로 2014년 1월에 높은 값을 보였으며 2013년 8월에는 매우 낮은 값을 나타냈다. 저서성 요각류의 계절별 서식밀도는 2014년 4월에 가장 높은 131 inds./10 cm2가 출현하였고, 2013년 8월에 가장 낮은 38 inds./10 cm2가 출현하였다. 갑각류유생의 계절별 서식밀도는 2014년 4월에 가장 높은 146 inds./10 cm2가 출현하였고, 2013년 8월에 가장 낮은 19 inds./10 ㎝²가 출현하였다. 편형동물의 계절별 서식밀도는 2014년 4월에 가장 높은 45 inds./10 cm2가 출현하였고, 2013년 8월에 가장 낮은 3 inds./10 cm2가 출현하였다(Table 2). 서해 및 유사한 퇴적물에 서식하는 중형저서동물에 대한 연구결과를 살펴보면 계절적으로 약간의 분류군 차이는 있지만 선충류, 저서성 요각류, 저서유공충류, 갑각류유생이 가장 우점하는 분류군으로 나타났다(Kim et al., 1998; Kim et al., 2004; Min et al., 2006; Kim et al., 2014).
출현 분류군 수
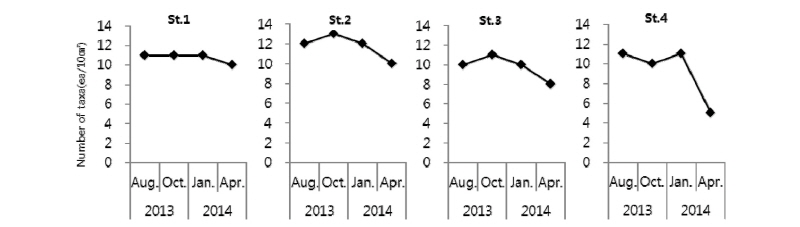
각 정점별‧계절별 중형저서동물 출현 분류군 수는 Fig. 4에 나타냈다. 가장 높은 중형저서동물 출현 분류군 수는 2013년 10월 St. 2에서 13개로 나타났고, 2014년 4월 St. 4에서 5개로 가장 낮게 나타났다. 각 계절별로 출현한 중형저서동물 분류군 수는 평균 8∼11개 분류군으로 나타났다. 각 정점별로 출현한 중형저서동물 분류군 수는 St. 2에서 일반적으로 가장 높게 나타났고, 그다음으로는 St. 1이었으며, St. 3, 4는 유사한 출현 분류군 수를 나타냈다. St. 4에서의 중형저서동물 출현 분류군 수는 2014년 1월에 비하여 4월에 급감하였는데, 이는 2014년 1월에 매우 낮은 서식밀도로 출현하였던 패충류(Ostracods), 이매패류(Bivalves), 단각류(Amphipods), 동문동물(Kinorhynchs) 등이 2014년 4월에는 전혀 출현하지 않았기 때문이다. 또한 출현 분류군수에 비하여 총 서식밀도가 높게 나타나는 이유는 우점종인 선충류의 서식밀도가 매우 높기 때문이다. 선충류는 각 정점의 대부분의 시기에서 약 90%이상의 비율을 나타냈다. 일반적으로 조간대의 세립질 퇴적물에서 선충류가 우점하는 경향이 많은 것으로 알려져 있으며(Giere, 1993), 본 연구지역에서도 모든 시기와 정점에서 가장 우점하는 분류군으로 나타났다. Kim et al.(1998)에 의해 새만금 갯벌에서 조사되어진 중형저서동물 군집 조성 결과에서도 총 12개의 분류군이 출현하였고, KORDI(2001)에 의해 대부도 갯벌에서 조사되어진 중형저서동물 군집 조성 결과에서도 총 13개의 분류군이 출현하였으며, 또한 Min et al.(2006)에 의해 태안 이원방조제에서 조사된 중형저서동물 군집 조성 결과에서도 출현한 중형저서동물 분류군 수는 12개로, 본 연구와 큰 차이를 보이지 않았다. 퇴적상이 사질형태인 장봉도 조간대에서 진행된 중형저서동물 군집 연구에서도 출현한 분류군 수가 11개로 본 연구와 큰 차이를 보이지 않았다(Kang et al., 2011).
중형저서동물 크기 분포
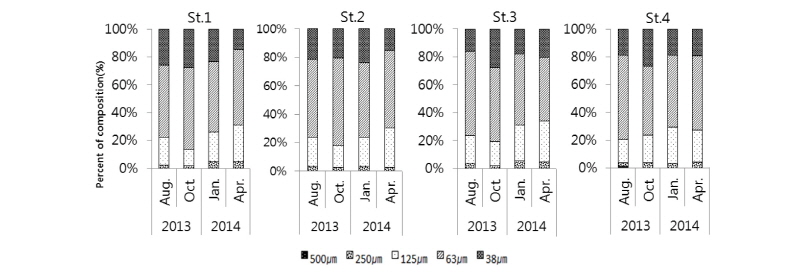
태안 근소만 지역에 서식하는 중형저서동물의 크기 분포는 모든 정점에서 63 µm ∼125 µm 사이 크기의 개체가 가장 많이 서식하는 것으로 나타났다(Fig. 5). 그다음으로는 38∼63 µm 사이 크기의 개체가 대부분의 정점에서 높게 나타났고 상대적으로 250 µm 이상 크기의 개체가 차지하는 비율은 매우 낮은 값을 나타냈다. 가장 높은 중형저서동물 크기 분포비를 갖는 63 µm∼125 µm 크기 개체들의 서식밀도는 정점별‧계절별로 살펴보면 다음과 같다. 2013년 8월에 St. 1에서 2,072 inds./10 cm2로 전체의 52%, St. 2에서 1,043 inds./10 cm2으로 전체의 55%, St. 3에서 1,076 inds./10 cm2으로 전체의 60%, St. 4에서 929 inds./10 cm2으로 전체의 61%의 비율을 차지하였다. 2013년 10월에는 St. 1에서 1,923 inds./10 cm2로 전체의 59%, St. 2에서 2,802 inds./10 cm2으로 전체의 62%, St. 3에서 1,640 inds./10 cm2으로 전체의 53%, St. 4에서 964 inds./10 cm2으로 전체의 50%의 비율을 차지하였다. 2014년 1월에는 St. 1에서 1,818 inds./10 cm2로 51%, St. 2에서 2,859 inds./10 cm2으로 전체의 53%, St. 3에서 2,191 inds./10 cm2으로 전체의 51%, St. 4에서 2,783 inds./10 cm2으로 전체의 52%의 비율을 차지하였다. 2014년 4월에는 St. 1에서 4,115 inds./10 cm2로 전체의 54%, St. 2에서 4,304 inds./10 cm2으로 전체의 55%, St. 3에서 2,240 inds./10 cm2으로 전체의 46%, St. 4에서 1,064 inds./10 cm2으로 전체의 54%의 비율을 차지하였다. 전체 중형저서동물 서식 밀도에서 63∼125 µm 크기 개체들이 차지하는 비율은 50% 이상을 나타내고 있어 본 연구에서는 크기가 작은 개체들이 많이 출현하는 것으로 나타났다. 일반적으로 크기가 작은 중형저서동물들은 큰 중형저서동물보다 생활사 기간이 상대적으로 짧고, 기회종으로 서식하는 경우도 많은 것으로 알려져 있다. 서해안 새만금 방제조 해역과 장봉도 해역에서 조사된 중형저서동물 군집 연구 결과에서 중형저서동물 크기 분포는 본 연구와 유사하게 63∼125 µm 사이 크기의 개체가 가장 많이 서식하는 것으로 나타났다(Min, 2007; Kang et al., 2011). 또한 기존 연구들에서 중형저서동물의 크기가 250 µm 이상의 개체는 전체 서식밀도의 2∼6% 비율로 나타나 본 연구결과인 2∼5%와 유사한 결과를 나타내었다(Min, 2007; Kang et al., 2011). 계절에 따른 중형저서동물의 크기별 분포의 차이에는 가장 우점하는 선충류(Nematodes)가 대부분의 기여를 하였고, 중형저서동물 분류군과 크기별 분포 차이가 특기할 만한 연관성을 나타내지는 않았으며 향후 선충류의 종 동정 등의 분석을 통하여 보다 확실한 결과를 도출할 필요가 있다고 사료된다.
생물량
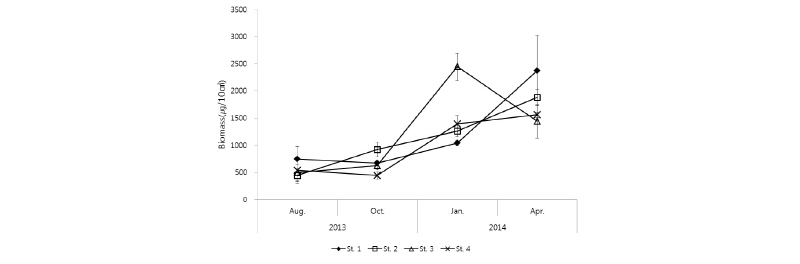
본 연구에서 조사 지역 내 전체 중형저서동물 생물량은 2013년 8월에 낮고, 2014년 4월에 높은 경향을 나타냈다(Fig. 6). 전체 중형저서동물의 평균 생물량은 556±238∼1,815±128 µg/10 cm2의 범위를 나타냈으며, 2014년 1월에 St. 3에서 2,446 µg/10 cm2으로 가장 높고, 2013년 8월에 St. 2에서 445 µg/10 cm2로 가장 낮은 값을 나타냈다(Fig. 6). 가장 높은 중형저서동물 생물량 값은 2014년 1월에 St. 3에서 나타냈는데, 이는 다른 분류군에 비해 상대적으로 생물량이 높은 저서성 요각류의 서식밀도가 급증하였기 때문이다. 각 정점의 2013년도 8월과 10월의 중형저서동물 생물량 값은 대부분 다른 조사시기보다 낮은 값으로 나타났고, 정점들 간에 큰 차이를 보이지 않았다. 중형저서동물 생물량은 모든 정점에서 2014년도 1월부터 서서히 증가하는 경향이 나타났다.
중형저서동물 생물량의 계절적 변동을 살펴보면, St. 1에서는 2013년 8월보다 10월에 소폭 감소하였다가 2014년 1월에 소폭으로 증가하였고 4월에는 크게 증가하였다. St. 2에서는 2013년 8월에서 2014년 4월까지 서서히 증가하는 양상을 보였다. St. 3에서는 2013년 10월에 약간 증가하였고, 2014년 1월에 큰 폭으로 증가하였다가 4월에 급격한 감소를 나타내어 대부분의 정점에서 2014년 1월에서 4월까지 생물량이 증가하는 값을 보인 것과는 다른 경향을 보였다. St. 4는 2013년 8월과 10월에 유사한 값을 나타냈고 2014년 1월에 급격하게 증가하였다가 4월에 소폭 증가하는 값을 보였다.
각 정점별로 중형저서동물 생물량 값을 살펴보면, St. 1에서 2014년 4월에 2,373±648 µg/10 cm2으로 가장 높고, 2013년 10월에 689±4 µg/10 cm2으로 가장 낮게 나타났다. St. 2에서는 2014년 4월에 1,884±135 µg/10 cm2으로 가장 높고, 2013년 8월에 445±118 µg/10 cm2으로 가장 낮게 나타났다. St. 3에서는 2014년 1월에 2,446±254 µg/10 cm2으로 가장 높고, 2013년 8월에 495±153 µg/10 cm2으로 가장 낮게 나타났다. St. 4에서는 2014년 4월에 1,559±52 µg/10 cm2로 가장 높고, 2013년 10월에 444±67 µg/10 cm2로 가장 낮게 나타났다. 서해 다른 연구지역에서의 중형저서동물 생물량 결과를 보면 계절별·정점별로 차이는 있으나 여름철보다 겨울철에 생물량 값이 낮은 경향을 보였다(Kang et al., 2011; Kim et al., 2014). 그러나 본 연구에서는 대부분의 정점에서 여름철인 2013년 8월에서 봄철인 2014년 4월로 갈수록 생물량 값이 증가하는 경향을 보였다. 또한 2014년 1월 St. 3에서 중형저서동물 서식밀도 변화와는 다르게 생물량이 급격하게 증가하였는데, 이러한 현상은 500 µm∼1 mm 사이 크기의 선충류(Nematodes)와 저서성 요각류(Harpacticoids) 비율이 다른 정점·계절에 비하여 상대적으로 매우 높게 나타났기 때문이다.
수직분포
각 정점의 0∼3 cm 퇴적물 내 중형저서동물 수직분포를 분석한 결과, 대부분 정점에서 퇴적물의 깊이가 증가함에 따라 중형저서동물 서식밀도가 감소하는 것으로 나타났다(Fig. 7). 이렇듯 퇴적물 표층으로부터 그 깊이가 깊어짐에 따라 중형저서동물의 서식밀도가 서서히 감소하는 것은 먹이나 산소 조건 등이 퇴적물의 깊이에 따라 열악해지기 때문에 나타나는 가장 일반적인 경향이라 할 수 있다(Higgins and Thiel, 1988). 중형저서동물의 수직분포 특성은 먹이의 분포, 용존 산소량, 공극률, 함수율 등이 분포를 결정짓는 주요 환경적 요인으로 알려져 있다(Dye, 1983; Jiggins and Thiel, 1988; Giere, 1993; Danovaro, 1996; Gwyther and Fairwather, 2005).
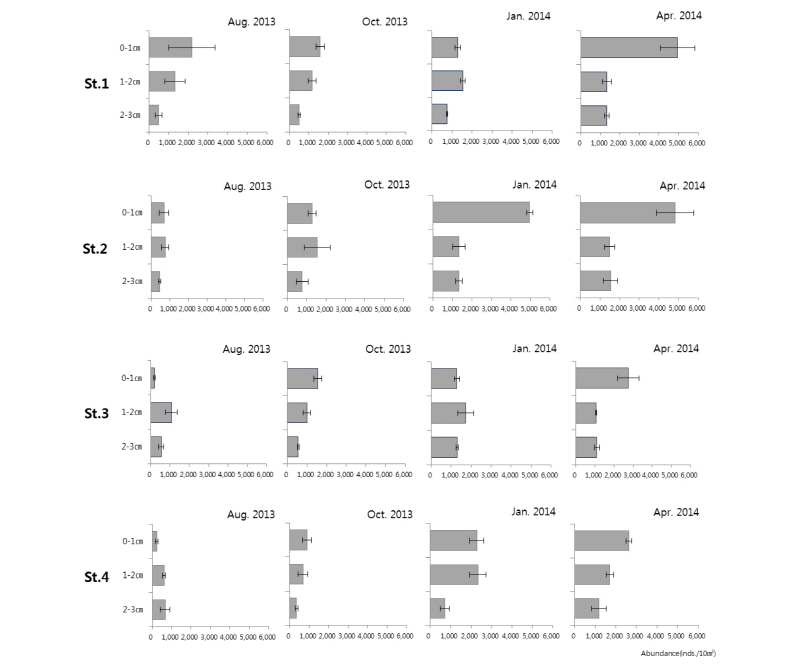
2014년 1월의 St. 2와 2014년 4월의 St. 1, 2에서의 중형저서동물 수직분포는 0∼1 cm 깊이에서 아주 높은 서식밀도를 보이고, 그 이하 깊이에서는 서식밀도가 급감하는 경향이 보였다. 2013년 8월의 St. 1, 10월의 St. 1, 3, 4 그리고 2014년 4월 St. 3, 4에서의 중형저서동물 수직분포는 표층에서부터 깊이가 깊어질수록 서식밀도가 감소하는 일반적인 중형저서동물 수직분포 형태가 보였다. 수직분포의 계절변동 값은 St. 1에서는 2013년의 8월과 10월, 2014년의 1월에 유사한 값의 서식밀도를 보였으나 2014년 4월에 표층의 서식밀도가 급증하는 결과를 나타내었고, St. 2에서는 2013년 8월과 10월의 서식밀도 값이 유사하나 2014년 1월과 4월에 표층의 서식밀도가 급증하는 결과를 나타내었다. St. 3에서는 2013년 8월에 표층의 서식밀도 값이 매우 낮았으나 2013년 10월, 2014년 1월에 증가하였고 2014년 4월에 가장 높은 값을 나타내었다. St. 4에서는 2013년 8월에 표층의 서식밀도 값이 매우 낮았으나 2013년 10월에 소폭 증가하였고, 2014년 1월을 지나 4월로 갈수록 증가하는 서식밀도 값을 나타내었다. Xiao-Shou Liu et al.(2007)에 의해 연구된 황해 중국연안의 중형저서동물 수직분포 특성에서도 전체 중형저서동물 서식밀도의 78.78%, 선충류의 76.17%와 저서성 요각류의 92.54%가 표층 2 cm 이내에서 출현하는 것으로 나타났다. 그러나 2013년 8월의 St. 3, 4, 2013년 10월의 St. 2 그리고 2014년 1월의 St.1, 3에서의 중형저서동물 수직분포는 일반적인 수직분포 특징과는 다르게 표층인 0∼1 cm보다 아래 깊이인 1∼2 cm에서 중형저서동물이 많이 서식하고 있는 결과를 보였다. 그러나 이들 정점도 대부분 2∼3 cm 깊이에서 서식밀도가 다시 급격히 감소하는 것을 보였다. 2013년 8월의 St. 3, 4에서 중형저서동물 수직분포는 표층 0∼1 cm 깊이의 서식밀도가 2∼3 cm 깊이의 서식밀도보다 매우 낮은 서식밀도 값을 나타냈다. 이러한 현상처럼 중형저서동물 군집이 산소공급이 원활하고 먹이조건이 좋은 표층을 선호하지 않은 이유로는 서식에 부적합한 요인이 발생하였다고 생각할 수 있다. 부적절한 요인으로는 인간에 의한 인위적인 교란이나, St. 3, 4가 조간대 상부이기 때문에 조간대 중하부에 비하여 한여름에 태양에 노출되는 시간이 길어져서 표층의 온도가 높아졌기 때문에 표층 이하로 내려가 있을 가능성이 있다. 이러한 현상은 일반적인 중형저서동물의 수직분포 현상은 아니지만 일부 다른 연구에서도 발생하고 있었음을 알 수 있다(Min et al., 2006).
집괴 분석
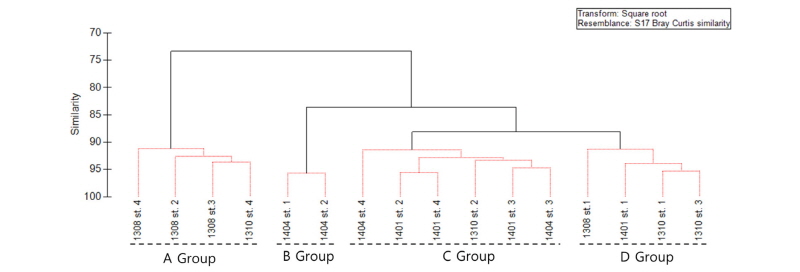
조사기간 동안 출현한 중형저서동물의 각 분류군과 개체수의 자료를 이용하여 집괴분석과 SIMPROF 분석을 실시하였다(SIMPROF test, P<0.05; Fig. 8). 집괴분석 결과 4개의 그룹으로 구분되어졌다. A 그룹에는 2013년 8월 St. 2, 3, 4와 2013년 10월 St. 4가 포함되고, B 그룹은 2014년 4월의 St. 1, 2를 포함하고, C 그룹은 2013년 10월의 St. 2와 2014년 1월의 St. 2, 3, 4, 2014년 4월의 3, 를 포함하고 있다. D 그룹에는 2013년 8월의 St.1, 2013년 10월의 St. 1, 3과 2014년 1월의 St. 1이 포함되어있다. A 그룹의 정점들은 대부분 기온이 높은 여름에 조사가 진행된 정점들이 포함되어있다. B 그룹에는 춘계 조간대 상부 정점들이 포함되어 있다. C 그룹에의 정점들은 기온이 낮은 겨울에 조사가 진행된 정점의 비율이 높았다. D 그룹에는 여름, 가을, 겨울에 조사된 St. 1이 포함되었다.
생물 지수
최근 중형저서동물을 이용한 환경 평가 방법에 대한 관심이 증가하고 있다(Sandulli and Be Nicola, 1990). 일반적으로 선충류는 오염, 특히 빈산소 환경에 대한 내성이 강하고 저서성 요각류는 오염에 민감한 생태적 특징을 가지고 있다(Raffaelli and Mason, 1981). 특히 저서성 요각류는 중형저서동물 분류군 내에서 다른 분류군들에 비하여 환경 변화에 민감하며(Hargrave and Thiel, 1938; Warwick and Clarke, 1991; Coull and Chandler, 1992), 연구 지역의 오염도와 비례하여 개체수의 급격한 차이를 보이는 대표적인 분류군으로 알려져 있다(Raffaelli, 1981, 1987; Sandulli and De Nicola, 1990). 이러한 특징을 이용하여 오염도를 평가하는 지수인 N/C ratio를 이용하여 연구지역 저서생태계의 건강도를 나타내었다. N/C ratio는 값이 높을수록 오염도가 증가하고, 값이 낮을수록 오염도의 감소를 지시한다.
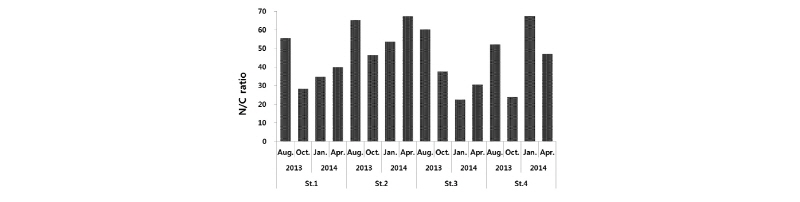
본 연구에서의 N/C ratio 값은 계절별로 평균 34∼58.8의 범위를 나타냈다. 가장 낮은 N/C ratio 값은 2014년 1월 St. 3에서 22.5로 나타났고, 2014년 1월 St. 4에서 67.5로 가장 높은 N/C ratio 값이 나타났다(Fig. 9). 새만금 외해역 지역의 연구에서의 N/C ratio 값이 0.3∼334.3의 넓은 범위를 보이는 것에 비하여(Kim et al., 2014), 본 연구에서는 N/C ratio 값의 범위가 22.2∼67.5로 비교적 좁은 범위의 값을 나타냈다. 하지만 광양만 지역에서의 평균 N/C ratio 값이 0.6∼0.83, 가만만 지역에서는 0.78∼0.9, 진해만 지역에서는 0.85∼0.9 값을 보였고(Lee, 2013), 이에 비하여 본 연구지역에서는 평균 N/C ratio 값이 34∼58.8으로, 상대적으로 넓은 범위의 값을 나타냈다. 연구지역 내의 각 계절별‧정점별 간의 오염도는 새만금 외해역 지역에 비하여 크게 차이가 나지 않지만 광양만, 가막만, 진해만 지역에 비해서는 정점 간의 차이가 다소 차이가 있다고 생각되어진다.
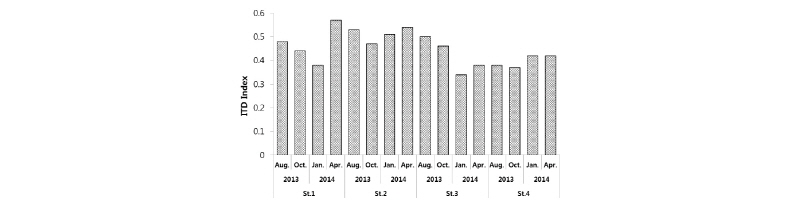
ITD 지수는 중형저서동물 그룹 중 가장 우점하는 선충류의 섭식 유형 4가지를 구분하고 이를 지수로 활용하여 값이 0.25∼1.0의 범위 값을 갖는다(Heip et al., 1985). ITD 지수는 값이 높으면 오염도가 증가하고, 값이 낮으면 오염도의 감소를 지시한다. ITD 지수를 이용하여 근소만의 계절별, 정점별 오염도를 나타낸 결과는 Fig. 10과 같다. 계절별 ITD 지수는 2014년 4월에 St. 1에서 값이 0.57으로 가장 높게 나타났고, 2014년 1월에 St. 3에서 0.34로 가장 낮게 나타나 2014년 1월에 오염도가 가장 높은 것으로 나타났다. 모든 정점에서 여름보다 가을에서 ITD 지수 값이 낮아졌고, 겨울에서 봄으로 가면서 ITD 지수 값이 높아지는 계절적 변동을 나타냈다. 이는 여름에서 가을이되며 오염도가 감소했다가, 겨울에서 봄이 되면서 오염도가 증가하는 경향을 나타내고 있다고 보여진다. Lee(2013)에 의하여 수행된 연구에서, 광양만과 진해만의 조사에서 2월에 비하여 8월에 ITD 지수 값이 높아져 오염의 증가를 지시했다는 결과와 일치하는 경향을 보인다. 또한 가막만, 광양만, 진해만에서 조사 정점이 하수처리장, 양식장 등의 육상 오염원 유입을 고려하여 청정지역과 오염지역으로 구분되었고, 가막만에서의 청정지역에서는 0.33, 오염지역에서는 0.68의 ITD 지수 값을 나타내었다. 광양만에서의 청정 지역은 2월에 0.42, 8월에 0.53의 값을 나타내었고, 오염지역에서 2월에 0.53, 8월에 0.56의 값을 나타내었고, 진해만의 청정지역에서는 2월에 0.38, 8월에 0.50의 값을, 오염지역에서는 2월에 0.43, 8월에 0.68의 ITD 지수 값을 나타내어 본 연구지역에서의 ITD 지수 값과 유사한 범위의 값을 보였다.
가장 오염도가 높게 나타난 2014년 4월의 St. 1에서는 선택성퇴적물식자(1A, Selective deposit feeder)가 우점하여 나타났고, 가장 오염도가 낮게 나타난 2014년 1월의 St. 3에서 출현한 선충류 그룹은 비선택성퇴적물식자(1B, Non-selective deposit feeder)가 가장 우점하여 나타났으며, 2014년 4월의 St. 1에서 선택성 퇴적물식자가 매우 높은 값으로 편향되어 나타난 것에 비하여 다양한 섭식 유형을 갖는 선충류(Nematodes) 그룹이 출현하였다. 즉, 이러한 현상으로 언급할 수 있는 것은 오염도가 높은 2014년 4월의 St. 1의 경우 선택성퇴적물식자가 상대적으로 높은 서식밀도를 보였고, 선택성퇴적물식자에 해당하는 선충류의 종이 특정 오염물질을 먹이로 섭취하고 있을 가능성이 있다는 것을 시사해 준다는 것이다. Lee(2013)의 연구에 의하면 광양만, 가막만, 진해만에서 각 만의 청정지역에서는 다양한 섭식 유형을 갖는 선충류가 출현하였고, 반면 오염지역에서는 퇴적물 섭식 유형의 선충류만이 출현하여 오염지역은 선충류 중에서도 특정한 그룹만이 서식할 수 있는 환경조건일 가능성이 있다는 결과를 나타내어 본 연구와 유사한 경향을 보여 주고 있다. 하지만 이러한 결과 값을 명확히 밝히기 위해서는, 향후 해당 시기의 해당 정점에서의 선충류 우점종에 대한 위 내용물 분석 등을 통하여 보다 확실한 연구 결과를 도출할 필요가 있을 것이다.
오염도가 가장 낮게 나타난 지역은 N/C ratio와 ITD 지수 모두 2014년 1월의 St. 3으로 나타났고, 오염도가 가장 높은 지역은 N/C ratio는 2014년 1월의 St. 4, ITD 지수는 2014년 4월의 St. 1로 두 값이 다른 결과를 보였다. 하지만 N/C ratio는 항상 오염에 비례하는 것은 아니며, 특히 물리적 환경의 변화가 이루어지는 지역에서는 큰 의미를 갖기는 어렵다(Back et al., 2009). 유기물 함량 등의 다른 요소와 비교해보면 N/C ratio가 근소만 지역에서 유효한 결과를 나타냈는지에 대하여 정확히 알 수 있으나 본 연구에서는 환경 자료가 부족하여 결론을 내리기에는 어려움이 있다. 하지만 생물지수를 적용한 연구는 국내 연안지역에서 아직 매우 부족한 실정이기 때문에 향후 환경 자료를 포함하여 생물지수의 유효성에 대한 추가적인 연구가 필요하다고 생각된다.
중형저서동물 군집의 계절별 특성을 파악하기 위해서는 많은 환경 자료와 장기간의 군집 모니터링이 요구된다. 본 연구에서는 1년 동안 4계절의 중형저서동물 군집 변화를 연구하였고, 환경 자료가 많이 부족한 관계로 환경 인자와의 상관관계를 파악하기에는 부족하나 향후 국내 조간대 갯벌에 서식하는 중형저서동물 군집의 생태학적 역할 등을 체계적으로 연구하는 데 있어서 중요한 기초자료가 될 것으로 사료된다.
