

서 론
재료 및 방법
Alexandrium 종주 확보
- 시료 채집
- 순수 분리
- 배양
Alexandrium 형태 및 유전학적 특성
- 형태적 특성 분석
- 유전학적 특성 분석
- 계통 분류학적 분석(Phylogenetic analysis)
결 과
종주별 형태학적 특성
Alexandrium sp. strain WEB-Alex-01
Alexandriumaffine strain WEB-Alex-02
Alexandriumcatenella strain WEB-Alex-03
Alexandrium fraterculus strain WEB-Alex-04
종주별 DNA 염기서열 유사도 분석 결과
Alexandrium sp. strain WEB-Alex-01
Alexandriumaffine strain WEB-Alex-02
Alexandriumcatenella strain WEB-Alex-03
Alexandrium fraterculus strain WEB-Alex-04
분자계통학적 특성
토 의
요약 및 결론
1. 서 론
해양생태계에서 발생하는 다양한 현상 중에 하나인 ‘적조(red tides)’는 플랑크톤 등이 일시에 다량으로 증식되거나, 물리학 작용으로 일정 지역에 집적되어 해수의 색을 변색시키는 현상을 말한다(ECOHAB, 1995). 이러한 적조 생물의 증식과 집적은 수중의 용존 산소를 고갈시켜 다른 생물의 생존을 저해하며(Hallegraeff, 1993), 규조류와 더불어 적조원인의 주된 그룹을 형성하는 와편모류의 일부는 독성을 함유하고 있어, 해양 먹이망을 교란시키고, 어패류를 독화 시키는 등의 생태계 균형 붕괴의 원인이 된다. 또한 이러한 독성을 함유한 유해 미소생물로 인해 독화된 패류를 사람이 섭취하였을 때, 사람의 건강까지도 위협을 주고 있기 때문에 사회‧경제적으로 심각한 문제가 되고 있다(ECOHAB, 1995; Anderson, 1997; Park et al., 2013).
해양 생태계에서 발생하는 독성으로 가장 널리 알려진 마비성패독(paralytic shellfish poisoning, 이하 PSP)은 ‘saxitoxin(또는 STX)’이라는 높은 수치의 수용성 신경독으로, 주로 독성 와편모류에 의해 독화된 패류를 섭식함으로 두통과 신체의 마비, 호흡곤란 심지어 사망의 원인이 되는 심각한 해양생물기원의 유해 물질이다(Kao, 1993; Bricelj and Shumway, 1998; Park, 2008). 국내의 경우, 1986년 3월 부산 감천만에서 진주담치를 섭식하고 2명이 사망하는 사고가 공식적으로 최초 보고 되었고(Chang et al., 1987), 1996년 5월 경남 거제군 장목면 인근 갯바위에서 채집한 진주담치를 끓여 먹고 3명이 중독되어 이 중 2명이 사망하는 사고가 발생하기도 하여(Lee et al., 1997), 이미 1987년부터 PSP에 대한 모니터링이 국립수산과학원 주도로 이루어지고 있다.
해양 와편모류인 Alexandrium 속에 속하는 종들은 가장 널리 알려진 PSP 원인 생물로서, 전세계 기수역 또는 연안역에서 출현하고, 아북극 지역, 온대 그리고 열대 지역 등 전 세계 거의 모든 해역에서의 PSP 발생원인 종이다(Taylor et al., 1995). Alexandrium 속에 대한 최초 연구는 1904년 Paulsen에 의해 Goniodoma ostenfeldii로 명명된 종을 대상으로 연구를 시작하여 2015년 현재, 약 33종이 학계에 보고되어 있으며, type species는 Alexnadrium minutum Halim(1960)이다. 전 세계적으로 Alexandrium의 출현과 이로 인한 PSP 검출 동태에 관한 연구에 의하면, 1970년대에는 미국, 유럽, 일본, 호주 등을 중심으로 한 출현보고가 우세하였으나(Delgado et al., 1990; Giacobbe et al., 1996; Anderson, 1997; Wyatt and Jenkinson, 1997; Townsend et al., 2001) 2000년대 이후 아프리카, 아시아, 남미 등 전 세계로 확장되고 있는 추세이며(Wang et al., 2006; Lilly et al., 2007; Bolch and Salas, 2007; Anderson, 2009; Anderson et al., 2012), 이러한 Alexandrium의 세계적 분포는 휴면포자(resting cyst) 단계를 가지는 생활사 특성(Reviewed by Wyatt and Jenkinson, 1997; Anderson et al., 2012)과 점차 증가되는 선박평형수(Ballast water)의 이동범위 및 이용량과 상당한 연관이 있는 것으로 보고되고 있다(Reviewed by Bolch and Salas, 2007).
그동안 Alexandrium 속에 대한 생리/생태적 연구는 이들이 전적으로 광합성 생물이라는 전제하에, 온도, 염분, 영양염 등에 대한 성장 반응 연구와 독성학적 연구, 생활사 연구 등이 주를 이루었지만(Montresor, 1995; Nagai et al., 2004; Lim and Ogata, 2005), 1990년대 들어 와편모류의 생존전략으로 혼합영양(mixo-trophy)에 대한 연구 결과가 속속 보고되면서, Alexandrium 속에 속하는 종 중에, A. catenella, A. tamarense A. minutum 등은 광합성 뿐 아니라, 다른 생물을 포식하는 혼합영양성 생물이며, 이러한 영양학적 전략은 이들의 개체군 동태 및 적조발생의 한 수단으로 작용할 가능성이 대두되기도 하였다(Nygaard and Tobiesen, 1993; Jeong et al., 2005a). 이러한 혼합영양성 미소 생물은 해양 미소생태계에서 1차 생산자임과 동시에 1차 소비자(포식자)로서, 종속영양성 박테리아(Seong et al., 2006), 남조류(Jeong et al., 2005b; Glibert et al., 2009), 미소-소형 편모충류(Stoecker et al., 1997; Li et al., 2000; Berge et al., 2008 ), 규조류(Bockatahler and Coats, 1993; Yoo et al., 2009), 그리고 다른 혼합성 와편모류(Jeong et al., 1999, 2005a, 2005c) 등과 같이 다양한 먹이를 섭식하고, 이와는 반대로 다른 혼합영양성 와편모류(Jeong et al., 2005a)와 다양한 종속영양성 와편모류(Jacobson and Anderson, 1996; Menden-Deuer et al., 2005), 그리고 섬모충류(Kamiyama and Matsutama, 2005) 및 후생동물플랑크톤(Stoecker and Sanders, 1985)의 먹이가 되기도 하여, 해양 생태계 먹이망 구조 와 물질 순환이해의 새로운 개념의 정립이 요구된다.
국내 연안에서 Alexandrium 속의 최초 출현보고는 1978년인 것으로 알려져 있으며(Cho, 1978), 1977년 7월 진해만에서 발생한 적조현상에 대해 보고한 자료를 보면 적조를 일으킨 종이 Gonyaulax sp.일 것으로 보고하였으나, 군체 형성(Chain-forming) 특성과 형태적 특성을 고려하여 후속 연구자에 의해 Alexandrium일 가능성이 높은 것으로 밝혀졌다. 그 이후 Alexandrium에 의한 적조 발생 및 출현동태 연구가 꾸준히 진행되어(Kim et al., 1990; Han et al., 1993; Kim, 1994; Lee et al., 2003; Park and Kang, 2009; Oh et al., 2012), 2014년 이전까지 국내 연안에서 출현하는 Alexandrium 속의 종들은 A. affine, A. catenella, A. fraterculus, A. minutum, A. tamarense 등 5종 정도인 것으로 알려졌다. 하지만 선박평형수의 사용 증가, 기후변화 등의 다양한 요인으로 신규로 국내 연안에 출현하는 해양 와편모류가 점차 증가하는 경향에 비추어 보면, 새로운 Alexandrium 종들의 출현을 간과할 수 없는 실정이다. 실제로, Shin et al.(2014)은 진해-마산만 퇴적물에서 2011년-2012년 발견한 Alexandrium cyst를 발아시켜 유영 세포(또는 영양 세포)로 배양에 성공한 후, 형태적, 유전학적 정보 분석을 통해 A. insuetum으로 판명된 연구 결과를 발표한 바 있으며, 2015년에는 포항 인근 해역에서 Alexandrium 신종(new species)의 출현 보고가 있었다(Lim et al., 2015).
본 연구에서는 국내 연안에 출현하는 Alexandrium 속의 종 다양성을 확인하고, 국내 연안의 신규 출현 종 여부를 확인하기 위하여 광범위한 현장 시료 채집을 통해 Alexandrium 종주를 확보하여 배양체를 확립하였고, 형태 및 분자생물학적 특성에 기초하여 종 분류를 실시하였다. 본 연구를 통해 최근 국내 출현 Alexandrium 종의 다양성을 확인하고, Alexandrium에 대한 지속적 연구 기반을 다져, 미래에 발생 가능한 피해에 대한 사전 정보를 확보하며, 유해 해양생물에 대한 신속 종주 탐색 기술(DNA probe 등) 개발을 위한 기초 자료를 제공하고자 한다.
재료 및 방법
Alexandrium 종주 확보
- 시료 채집
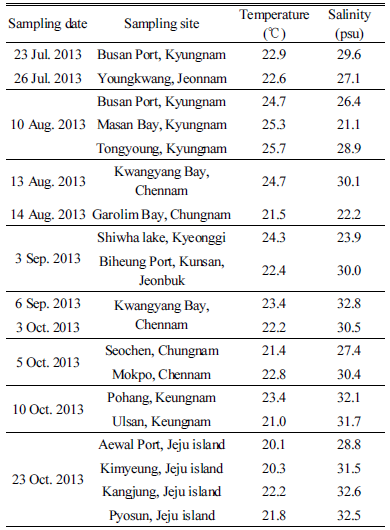
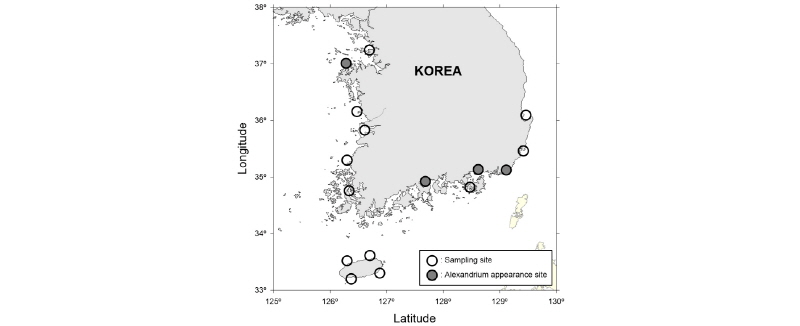
Alexandrium 종주 확보를 위해 2013년 7월부터 2013년 10월까지 전국 연안 지역을 대상으로 현장조사를 실시하였다(Fig. 1, Table 1). 정량 시료는 해수를 직접 또는 bucket을 이용하여 500 ml 폴리에틸렌 병에 채집하였고, 정성 시료는 식물플랑크톤 네트(망구 30 cm, 망목 : 20 µm)로 일정량 견인하여 농축된 시료를 채집하였다. 현장 시료 채집시 수온과 염분 특성은 YSI-1500을 이용하여 동시 측정하였고, 현장에서의 출현 농도를 파악하기 위해 500 ml 시료는 Lugol’s solution으로 고정하였다.
- 순수 분리
채집된 시료는 가급적 신속하게 실험실로 운반하여 즉시 순수 분리를 위한 처리를 실시하였다. 순수 분리는 해부 현미경 (Olympus SZX12, Olympus, Japan) 하에서 Alexandrium으로 예상되는 세포를 탐색하고, pasteur micro-pipette을 이용하여 단세포 또는 분열중 세포를 직접 포착하여 분리하였다. 분리한 세포는 무균상태의 해수(sterile seawater)에서 5회 이상 헹구어 준 뒤, 염분 28–30 psu의 f/2(-silicate) medi a를 채운 24 well culture plate(SPL lifesciences, Gyeonggido, Korea)에 옮겨 주었다.
- 배양
배양은 온도 20 °C(± 약 1.5 °C), 광도 20 µmol photons m-2s-1(연속광주기) 조건에서, Alexandrium 세포의 농도가 높아짐에 따라, 점차적으로 멸균된 f/2(-silicate) media를 넣은 6 well culture plate, 150 ml, 500 ml, 그리고 최종적으로 2 L polycarbonate bottles 순으로 계대 배양하였다.
Alexandrium 형태 및 유전학적 특성
- 형태적 특성 분석
최소 2 L 이상의 배양체가 확보되면 이들의 형태학적 특성 파악을 위해 먼저 현미경 관찰을 실시하였다. 살아있는 세포의 크기(길이와 너비) 측정과 전체적인 형태 특징은 광학 현미경(Zeiss-Axiovert 200 M, Carl Zeiss Ltd,. Göttingen, Germany)을 이용하여 400-1,000 배로 DIC(differential interference contrast illumination) 또는 bright field에서 관찰하였다. 정확한 종의 동정을 위해 먼저 와편모류의 thecal plates pattern의 관찰을 쉽고 빠르게 할 수 있는 calcofluor-white 염색법(Fritz and Triemer, 1985)을 사용하였다. 세포는 2%의 glutaraldehyde로 고정한 뒤, calcofluor-white M2R (Polysciences, Warrington, PA, USA)을 첨가하여 착색시키고 UV 필터장치(Zeiss HBO100, Carl Zeiss Ltd., Göttingen, Germany)가 설치된 도립형광현미경(Zeiss Axiovert 200 M, Carl Zeiss Ltd., Göttingen, Germany)으로 관찰하였다(excitation 330-385 nm, emission >420 nm). 일부 Alexandrium 종은 주사전자현미경(SEM)을 통한 세부 형태적 특성 분석도 병행하여 실시하였다. SEM 분석을 위한 시료는 2% glutaraldehyde로 고정시킨 후 osmic acid(OsO4)의 최종농도가 1-2%가 되도록 부가하여 실내에서 2시간 동안 고정하였다(Reimann et al., 1980). 이후 ethanol series로 탈수하여 isoamylace-tate로 치환시켜 Critical Point Dryer(SPI, Dry-CPD)로 건조시켰다. 건조된 시료는 aluminum stub에 고정하고 gold-palladium으로 코팅(JEOL, MSC-101) 후 주사전자현미경(SEM, JEOL 5600LV; 군산대학교 공동실험실습관 전자현미경실)으로 검경 및 동정하였다.
- 유전학적 특성 분석
일차적으로 현미경 관찰을 통해 Alexandrium 종으로 환인된 종들의 유전학적 특성 분석을 위해 ribosomal DNA의 Large subunit (LSU)구간의 분석을 다음과 같은 절차를 거쳐 실시하였다.
고농도(약 2,000 cells ml-1 이상)의 Alexandrium 배양체를 20 ml tube에 옮기고, 원심분리기(Micro 17TR, Hanil Science Industrial Co., Ltd., Incheon, Korea)를 이용하여, 실내온도에서 1,610×g의 속도로 10분 동안 농축시킨 후, 상층액은 버리고 농축된 시료(Pallets)는 1.5 ml tube로 옮겼다. 1.5 ml tube에 Proteinase K 20 µl, 전처리 용액 200 µl와 완충용액(lysis buffer) 200 µl를 넣고 5초 동안 섞은 후 항온 수조에서 60 °C로 1시간 동안 반응시킨 후, tube에 동량의 PCI-9(phenol chloroform isoamyl alcohol 25:24:1)을 넣고 5분간 1,610×g의 속도로 원심분리한 후 상층액을 새로운 1.5 ml 원심분리 tube로 옮겼다. 상층액에 2M sodium acetate 30 µl와 차가운 99.9% ethanol 900 µl를 넣은 후 가볍게 섞어서 DNA의 수율을 최대화하기 위해 -20 °C에서 30분 동안 두었다. 그 후 1,610×g의 속도로 5분 동안 원심분리한 후 상층액을 제거하고, 70% ethanol 500 µl를 넣고 가볍게 섞어준 뒤 1,610×g의 속도로 5분간 원심분리 시킨 후 상층액을 제거하였다. Ethanol을 완전히 제거하기 위해 오븐에서 1시간 동안 건조시킨 후 30 µl의 elution buffer를 첨가하여 DNA를 분리하였으며, 곧바로 실험에 사용 시엔 4 °C에 보관하였고, 장기간 보존하기 위한 시료는 -20 °C에 저장하였다. DNA 증폭은 forward primer: Euk1209F(5’-CAG GTC TGT GAT GCC C-3’), reverse primer: LSUB(5’-ACG AAC GAT TTG CAC GTC AG-3’)를 이용하였다(Medlin et al. 1988). PCR 반응액은 1×완충용액, 1.5 mM MgCl2, 0.2 mM dNTPs, 0.1% BSA, 5% DMSO, 2.5U Ex Taq polymerase(Takara), 0.5μM primer와 1 µl의 주형을 포함하여 최종 50 µl가 되도록 하였다. PCR 수행은 Eppendorf Mastercycler ep gradient (Eppendorf, New York, USA)를 사용하였고, pre-denature는 95 °C로 2분, denature는 95 °C로 20초, annealing은 55 °C에서 1분, elongation은 72 °C로 1분, post-elongation은 72 °C로 5분간 수행하였다. 증폭반응은 30회 반복 수행하였다. PCR 산물을 1% agarose gel에 전개하고 EtBr로 염색하여 UV하에서 DNA밴드를 확인하였다. LSU rDNA의 염기서열은 ABI PRISMⓇ 3700 DNA Analyzer(Applied Biosystems, Foster City, CA)를 이용하여 얻었다. 모든 염기서열은 ConfigExpress alignment program(InforMax, Frederick, MD)를 이용하여 분석하였다. 결정된 염기서열은 NCBI(National Center for Biotechnology Information)의 BLAST 검색(http://www.ncbi.nlm.-nih.gov)을 통해 유사도 분석(similarity analysis)을 실시하였다.
- 계통 분류학적 분석(Phylogenetic analysis)
유전학적 특성 파악에서 얻은 Alexandrium 종들에 대한 염기서열을 이용하여 각 종의 계통 분류학적 위치를 확인하였다. Alexandrium 종주들의 계통수는 Baysian(Huelsenbech and Ronquist, 2001) 및 Maximum Likelyhood(ML, Stamatakis, 2006) 방식을 이용하여 제작하였으며, 염기서열의 비교를 위해서 각 계통수에 사용된 염기서열은 NCBI에서 획득하여 이용하였다(Table 2). LSU rDNA의 염기서열 분석을 위해서는 MEGA v.4(Tamura et al., 2007)와 Clustal X2(Larkin et al., 2007)를 이용하여 분석 및 각 염기서열의 비교, 그리고 계통수 제작을 위한 염기서열의 정리를 실행하였다. 각 계통수의 ML 수치는 PAUP* 4.0b(Swofford, 2002)에서 제공되는 GTR+G+I 모델을 이용하여 제작하였다. Likelyhood 수치는 heuristic search 방식에 따라 100 random additional sequence replicate와 TBR branch swapping을 이용하여 분석되었으며 ML Bootstrap 수치 역시 획득되었다. Baysian 방식을 이용한 Bayesian consensus tree는 MrBayes v.3.1(Huelsenbeck and Ronquist, 2001; Ronquist and Huelsenbeck, 2003)을 이용하여 제작되었으며, ML tree와 마찬가지로 GTR+G+I 모델을 사용하였고, 이를 이용하여 얻은 Markov Chain Monte Carlo(MCMC)의 결과를 이용하여 substitution rates와 rate heterogeneity value(alpha), 그리고 base frequencies를 측정하였다. 각 계통수의 수치는 posterior probabilities ≥0.5를 기준으로 입력하였다.

결 과
한국 연안의 하계 Alexandrium 출현
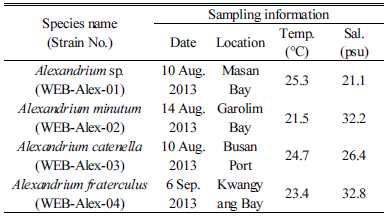
2013년 7월부터 10월까지 국내 16개 연안 표층에서 채집한 19개 식물플랑크톤 농축 시료 중에 마산만(strain WEB-Alex-01), 태안 가로림만(strain WEB-Alex-02), 부산항(strain WEB-Alex-03), 광양만(strain WEB-Alex-04) 해역에서 순수 분리 및 배양체 확립에 성공하여(Fig. 1, Table 3). 형태학 및 유전학적 특성을 분석하였다. 그 결과, 4개 종주는 각각 A. affine, A. catenella, A. fraterculus, 미동정(unidentified) Alexandrium sp.(strain WEB-Alex-01)로 확인되었고, A. catenella, A. affine, unidentified Alexandrium sp.(WEB-Alex-01)은 8월에, A. fraterculus는 이른 9월에 출현하였다. 각 지역별 출현 종주에 대한 형태적, 유전학적 특성에 대한 자세한 결과는 다음과 같다.
Table 3. Alexandrium strains. Species name, strain number, and origin of each strain such as date and location for water sampling as well as temperature and salinity of the water sample |
|
종주별 형태학적 특성
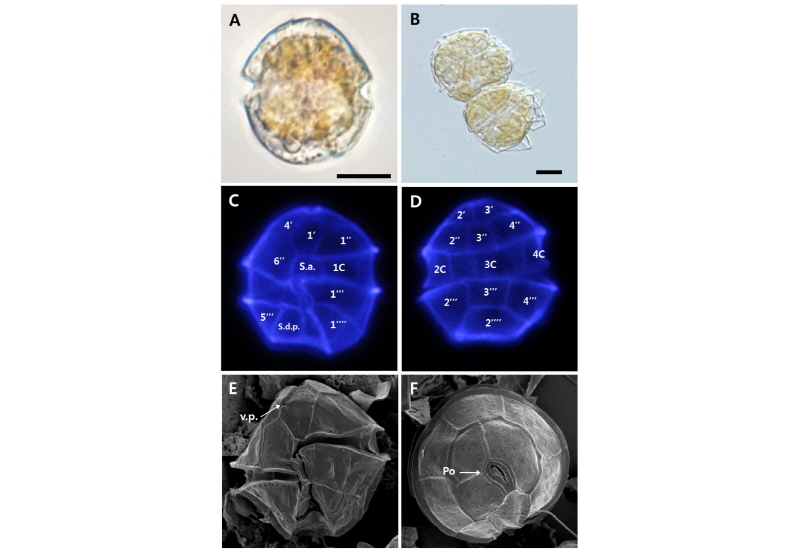
Alexandrium sp. strain WEB-Alex-01
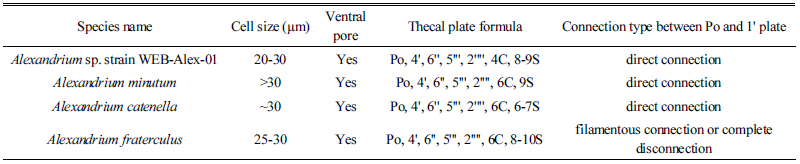
세포의 모양과 크기의 변이는 일부 있었지만, 전체적인 크기는 20-30 µm, 형태는 둥근모양이나, 세포의 외관이 매끈한 원형이기 보다는 울퉁불퉁한 특징을 가지며, 상각(epitheca)과 하각(hypotheca)의 높이는 거의 동일하였다. 상각은 볼록한 모양의 반구형이며, 하각의 모양은 비대칭이다. 세포 내부는 엽록소가 고르게 분포하는 특징을 보이고, 세포갑판(thecal plates)은 얇고, 쉽게 깨지는 성질이다. 세포의 세로판 홈(sulcus)은 세포 중간에서 하부로 내려갈수록 점차 확장되는 특징을 다수의 세포에서 확인할 수 있었다. Alexandrium 속의 분류에 있어서 중요한 3가지 갑판 분류키(key diagnostic plates) 가운데 하나인 ventral pore(v.p.)의 유무와 위치 특성을 확인해 본 결과, first apical plate(1')와 4' plate 사이 또는 1' 쪽으로 만입되어 있다. Kofoidian series에 준한 형태적 특성(thecal plate formula) 확인 결과, Po(apical pore plate), 4', 6'', 5''', 2'''', 4C, 8-9S로 구성되어 있으며, apical plate 1'는 Po와 직접적으로 이어져 있는(direct connection type) 경우가 대부분이었다. apical plate 3'과 4'은 5각형, 2'은 6각형이고, apical plates 중에서 가장 크기가 크다. Sulcal plates는 s.a., s.a.c., s.m.a., s.d.a., s.m.p., s.s.a., s.d.p., s.s.p., s.p.로 이루어져 있다. Antapical plate 1''''와 2''''는 오각형이며, 2''''의 크기가 더 크다. 이는 전형적인 Alexandrium 속에 해당하는 thecal plate formula 특징이며(Balech, 1995; Lilly et al., 2005), 그중에서도 Alexandrium minutum과 상당한 유사성을 보인다(Table 4, Fig. 2).
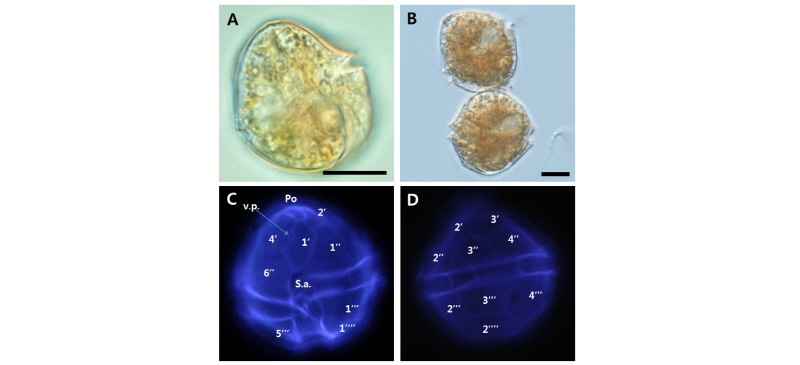
Alexandrium affine strain WEB-Alex-02
세포의 모양과 크기의 변이는 일부 있었지만, 전체적인 크기는 30 µm 이상이고, 형태는 오각형에 가까운 둥근(pentagonal round) 모양이며, 상각과 하각의 높이는 거의 동일하였다. 상각은 볼록한 모양의 반구형이며, 하각은 하단이 비교적 평평한 비대칭 구조이다. Kofoidian series에 의한 thecal plate formula는 Po, 4', 6'', 5''', 2'''', 6 C 그리고 9 S로 이루어져 있고, apical plate 1'는 Po와 직접적으로 이어져 있으며, v.p.는 일부 세포에서 확인되는데, 주요 위치는 4'와 만나는 지점의 중간 부근으로, 전형적인 Alexandrium 속의 thecal plate formula를 따랐다(Table 4, Fig. 3).
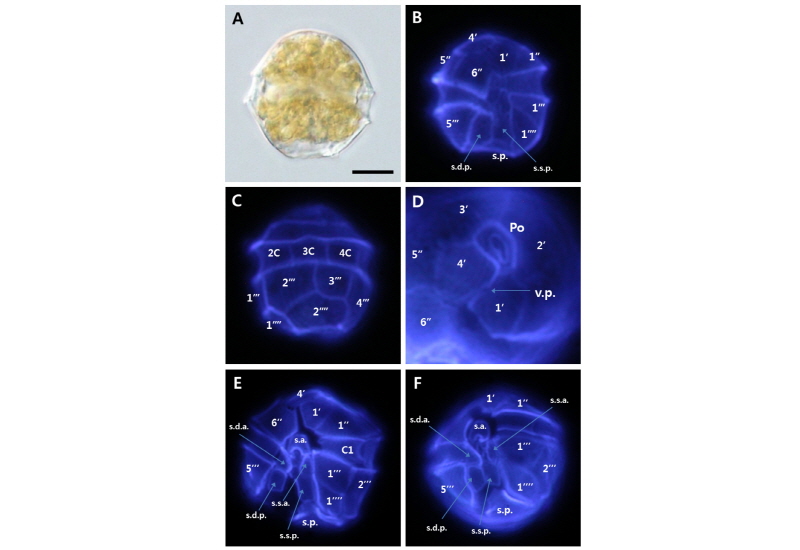
Alexandrium catenella strain WEB-Alex-03
전체적인 세포의 크기는 30 µm 내외이고, 형태는 둥근 모양, 상각과 하각의 높이는 거의 동일하였다. 상각은 볼록한 모양의 반구형이고, 하각의 하부는 sulcus의 두드러진 특징으로 오목한 비대칭 모양이다. thecal plate formula는 Po, 4', 6'', 5''', 2'''', 6C 그리고 6S 또는 7S로 이루어져 있으며, apical plate 1'는 Po와 직접적으로 이어져 있다. apical plate 4'은 2'와 연결되어져 있지만, precingular plate 1''과는 연결되어 있지 않았다. 3', 4', 5'은 5각형이며, 2'은 apical plates 중에서 가장 크다. Antapical plate 1''''와 2''''는 오각형이며, 2''''의 크기가 더 크다(Table 4, Fig. 4).
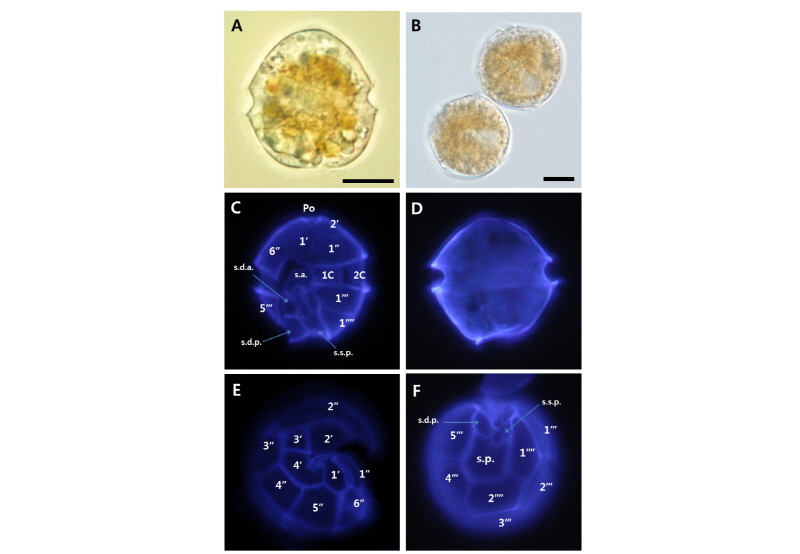
Alexandrium fraterculus strain WEB-Alex-04
세포의 전체적인 크기는 25-30 µm, 형태는 둥근 모양이고, 폭에 비하여 높이가 약간 큰 특징을 보이며, 상각과 하각의 높이는 거의 동일하였다. 세포 내부에는 많은 엽록소로 채워져 있고, 세포 갑판(thecal plates)은 얇고, 깨지기 쉬운 외부의 thecal membrane으로 덮여있다. thecal plates pattern은 Po, 4', 6'', 5''', 2'''', 6 C 그리고 8-10S로 이루어져 있으며, apical plate 1'는 Po와 약간 또는 완전히 이격된(filamentous connection or complete disconnection) 특징을 보인다. apical plate 4'은 2'와 간접적으로 연결되어져 있지만, precingular plate 1''과는 연결되어 있지 않았다. 3'과 4'은 5각형이며, 2'은 6각형이고, apical plates 중에서 가장 크기가 크다(Table 4, Fig. 5).
종주별 DNA 염기서열 유사도 분석 결과
Alexandrium sp. strain WEB-Alex-01
각 종주별로 획득된 LSU rDNA 염기서열의 유사도 분석(similarity analysis) 결과, Alexandrium sp. strain WEB-Alex-01 종주와 가장 유사한 DNA 서열구조를 가진 종은 Alexandrium sp. strain D164C5, D164C6(NCBI Accession No. EU707481, EU707480)으로, 99.9% 일치하는 결과를 보였다. 그 외에 종(species) 수준까지 비교 가능한 종 중에서 가장 유사한 종은 A. tamarense(EU707459)이었으나 일치율은 93% 수준으로, Alexandrium sp. strain D164C5, D164C6과 가장 유사한 염기서열 특징을 보이는 것으로 확인할 수 있다.
Alexandrium affine strain WEB-Alex-02
가장 유사한 LSU rDNA 염기서열 구조를 가진 종은 Alexandrium affine 종(JF906996, JF521618, JF521617, JF521616, AY831409)이었으며, 본 종주와 거의 0.001% 미만의 차이를 보인다. 즉, WEB-Alex-02 종주는 분자계통학적으로 A. affine인 것으로 확인되었다.
Alexandrium catenella strain WEB-Alex-03
본 종주와 가장 유사한 DNA 서열구조를 가진 종은 Alexandrium tamarense 종(JF521641) 또는 A. catenella 종(DQ785885, DQ785886, DQ785887, AY347308)인 것으로 확인되었다. NCBI 상에 등록된 A. tamarense(JF521641)의 경우, 2011년 1월 5,290 bp 길이의 자료가 등록된 CCMP1493 strain이고, A. catenella(DQ785885, DQ785886, DQ785887, AY347308)의 경우, 2007년 5월 6,370 bp 길이의 자료로서, 모두 본 WEB-Alex-03 종주의 1,242 bp 구간에서의 염기서열 상이성이 전혀 없는 관계로 보다 확장된 염기서열 분석이 요구되었다. 이에 따라, internal transcribed spacer 구간(ITS1, 5.8S, ITS2)부분에 대한 추가적인 염기서열 분석을 실시하였고, 이를 바탕으로 NCBI의 BLAST 검색 결과, WEB-Alex-03 종주는 Alexandrium catenella인 것으로 확인되었다.
Alexandrium fraterculus strain WEB-Alex-04
WEB-Alex-04 strain은 Alexandrium fraterculus 종(JF521623, JF521622, AY438017)과 100% 일치하는 것으로 확인되었다.
분자계통학적 특성
본 연구를 통해 확보한 4종의 Alexandrium 종주의 ribosomal DNA의 LSU구간에 대하여 NCBI에서 얻은 Togula compacta(AY-568562)과 T. jolla(AY455680)의 염기서열을 outgroup으로 적용하여 계통분류학적 분석(계통수 제작)을 하였을 때, Alexandrium sp. strain WEB-Alex-01 종주는 다른 종에 비하여 A. minutum A. insuetum, A. cf. tamutum 등과 비교적 가까운 계통분류학적 특성을 보이나 동일 분지되는 종은 존재하지 않는 것으로 판명되었다. 즉, 현재까지 보고된 Alexandrium 속에 속하는 유전학적 특성이 동일하거나 거의 유사한 종이 없는 신종(new species)의 가능성이 높은 종이라 할 수 있다. WEB-Alex-01 strain과는 달리 WEB-Alex-02 strain은 A. affine과 같이 분지되었고, WEB-Alex-03 strain은 A. catenella와 WEB-Alex-04 strain은 A. fraterculus와 같이 분지되었으며, 이를 뒷받침해주는 ML bootstrap 수치 및 baysian 수치 역시 높은 지지율을 보였다(Fig. 6).

토 의
2013년 하계에 우리나라 전국 연안에서 순수 분리한 Alexandrium 종은 A. catenella, A. affine, A. fraterculus 그리고 종(species) 수준의 동정이 이루어지지 않은 Alexandrium sp. 등 총 4 종이었다. 출현 농도는 생시료를 농축하였을 때만 현미경 관찰이 가능한 수준으로, 모두 1 cells ml-1 이하의 출현량을 보였다. 과거에 우리나라 연안에서 이루어진 Alexandrium 출현 연구를 종합해 보면, 1993년 9월부터 1994년 7월까지의 진해만 4개 정점에서 실시한 Alexandrium 유영세포 분포조사(김, 1994)에서 가장 높은 농도의 출현은 3월(533 cells ml-1)인 것으로 보고하였다. 마산만과 진해만 일원에서 1996년과 1997년 봄철(3월-5월) 실시한 주별 조사(Lee et al., 2003)에서는 A. tamarense가 10,000 cells ml-1의 고농도로 출현하였고, 수온은 12-17 °C로 비교적 낮은 수온에서 출현하는 것으로 보고하였다. 2008년 2월부터 11월까지 진해만 10개 정점에서 specific real-time PCR probe를 이용한 A. minutum 월별 출현 연구결과(Park and Kang, 2009)에서는 총 100여 개의 해수 시료중 A. minutum DNA가 검출된 경우는 총 3개의 시료이었고, 출현시기는 5월과 6월, 그리고 검출량은 1-4 cells l-1의 저농도인 것으로 나타났다. 또한 2011년 1월부터 4월까지 진해만 15개 정점을 대상으로한 임 등(2011)의 연구에 의하면, Alexandrium spp.는 1월에는 거의 출현하지 않으며, 2월에 최대 70 cells l-1로 출현하였다가, 3월초에 최대 400 cells l-1의 농도로 증가하였는데, 이는 수온상승과 관련성이 있는 것으로 보고하였다. 마산만에서 2004년 7월부터 2006년 5월까지 실시한 장기간 정밀 모니터링 결과(Jeong et al., 2013) 중, Alexandrium spp.로 분류한 결과만 선별하여 분석하면, 마산만에서 Alexandrium 출현은 2004년 8월에 최대 85 cells ml-1의 일시적인 고농도 출현이 있었으나 적조발생 수준의 농도는 아니었고, 이후 2005년 춘계부터 2006년 춘계까지 비록 낮은 농도이긴 하지만 모든 계절에 걸쳐 출현하는 것으로 조사되었다. 비교적 가장 최근의 연구인 오 등(2012)의 마산만 연구에서 Alexandrium의 출현은 4월부터 7월까지이며, 5월과 6월에 각각 8 cells ml-1와 35 cells ml-1로 와편모류 출현량의 약 60% 이상을 차지하는 우점종인 것으로 보고하였다. 이상의 연구를 종합해 보면 우리나라 연안의 Alexandrium 출현은 남해안의 진해-마산만 일원에 한정적이다. 하지만 본 연구에서는 서해안인 태안지역에서 A. affine이 출현하였고, 남해안의 광양, 부산지역에서도 다양한 Alexandrium 종이 출현하였다. 출현 시기 역시 기존의 연구에서는 대부분 춘계에 한정적인 것으로 알려 졌으나 본 연구 결과, 하계에도 다양한 Alexandrium 종이 출현하고 있음을 확인하였다.
부산항에서 채집, 순수 분리한 A. catenella는 A. tamarense와 더불어 과거 국내 연안에서 적조를 발생한 사례가 많은 PSP 패류독화의 주요 원인종으로, 두 종은 형태 및 유전학적으로 매우 유사한 특성을 보이나, 독성학적으로는 일부 특이성을 보이는 종이다(Balech, 1995; Taylor et al., 1995; Kim et al., 2002; Anderson et al., 2012; Oh et al., 2012). 또한 A. catenella는 A. tamarense, A. fundyense와 더불어 ‘Alexandrium tamarense species complex’를 구성하는 종으로 형태적, 유전적 특성 변화가 크지 않아 종의 구분이 모호한 것으로 오랫동안 연구되어져 왔다(Scholin et al., 1994; Anderson et al., 1994, 2012; Lilly et al., 2007). 2014년 발표된 John et al.(2014a)의 연구에서, 기존에 알려진 A. australiense, A. catenella, A. fundyense, A. pacificum, A. mediterraneum, A. tamarense를 Alexandrium tamarense species complex 내에 5개의 clade로 새롭게 편입, 구분해야 한다는 분류체계에 대한 주장이 제기된 바 있고, 더욱 최근에는 이들의 새로운 분류 체계 주장에 문제가 있으며, 나아가 A. catenella의 속명을 변경해야 한다는 의견에 대한 반론 주장이 개진되는 등(John et al., 2014b; Fraga et al., 2015) Alexandrium 속에 포함되는 종들의 분류체계에 대한 논란은 현재 진행중이다. 이렇듯 분류체계가 완전하지 않은 현 상황에서, 본 연구는 신속한 종의 분류에 널리 활용되는 ribosomal DNA의 Large subunit 구간만의 분석을 통해 A. catenella 출현을 확인하고, 춘계에 한정적으로 출현하여 패류독화의 원인으로 인식하고 있던 A. catenella(또는 Alexandrium tamarense species complex)가 하계에도 출현함을 확인한 결과이다.
우리나라 서해안에 위치한 태안 연안(가로림만)에서 순수 분리된 Alexandrium affine은 1985년 Fukuyo et al.에 의해 Protogo-nyaulux인 것으로 일본 연안에서 보고된 이래, 베트남, 칠레, 중국 연안 등에서 출현하는 것으로 보고되었으며(Nguyen-Ngoc, 2004; Wang et al., 2006), 비록 낮은 농도이긴 하지만 Saxitoxin을 함유한 유해종으로 알려져 있다. 국내의 출현보고는 1986년 9월과 1987년 10월에 A. fraterculus와 혼합하여 남해안의 진동만에서 적조를 유발한 바 있다는 연구결과(Kim et al., 1990)가 있고, Lee et al.(1998)의 연구에서도 1996년 6월부터 1997년 5월까지 마산-진해만에서 거의 매월 출현하는 것으로 보고된 바 있다. 이후 A. affine을 한국 연안에서 채집 및 순수 분리하여 실험에 사용한 경우(Kim et al., 2004; Kim, 2006)와 휴면포자 출현 연구(Shin et al., 2010; 2012)를 통해 우리나라 남해안에서만 출현하는 종으로 예상되었지만, 본 연구를 통해 서해안인 태안 연안에서 하계에 출현하고 있음을 처음으로 확인하였다.
남해안의 광양만에서 순수 분리한 A. fraterculus는 Gonyaulax fratercula, Gessnerium fraterculum, Protogonyaulax fratercula 등의 이명으로 학계에 알려져 있다가, 1964년 Balech에 의해 Alexandrium 속으로 분류된 종으로서, 아직까지 독성의 유무는 알려지지 않았다(Anderson et al., 2012). 국내의 출현양상은 1986년 9월과 1987년 10월에 A. affine와 혼합하여 남해안의 진동만에서 적조를 유발한 바 있다는 연구결과(Kim et al., 1990)가 있고, 1996년 6월부터 1997년 8월까지 마산-진해만에서 출현한 연구결과가 있으며(Lee et al., 1998), 1997년 6월 진해만에서 순수 분리한 A. fraterculus 종주에 대한 유전학적 연구가 진행된 바 있다(Lee et al., 1998). 또한 2009년 경남 남해군 일대에서 발생한 적조 원인종으로서, 최대 출현 농도는 2009년 8월 13일 경남 남해의 앵강만 등지에서 최대 6,000 cells ml-1의 농도로 출현한 바 있는(국립수산과학원, 적조정보시스템) 유해종이다. 따라서 유사한 시기에 적조를 유발하는 Cochlodinium polykrikoides와의 생태학적 역할 비교, 동일시기 출현시 어류 치사 촉진 여부, 정확한 독성 함유 여부 등에 대한 추가적인 연구가 필요한 종으로 판단된다.
2013년 8월 마산만에서 순수 분리한 Alexandrium sp.(WEB-Alex-01 strain)은 유전학적 특성 분석에서 본 strain과 동일한 유전적 정보를 가진 종은 Alexandrium sp. strains(NCBI 등록 종 정보 ID: D163 C5, D164 C6)인 것으로 확인되었는데, McCauley et al.(2009)의 연구에 의하면, 이 두 개 strain은 A. minutum(Pacific clade)과 상당한 유전학적 유사성을 보이지만 100 % 일치하지는 않고, 신종의 가능성이 높으나 종주의 배양체 확립에 실패하여 추가적인 형태적 특성 정보는 알 수 없다고 언급하고 있다. 또한 Shin et al.(2014)의 연구에서 진해-마산만의 퇴적물 내의 Alexandrium cyst를 발아시켜 배양에 성공한 A. insuetum 종과도 상당한 형태적 차이를 보이는데, 그들의 연구에서 A. insuetum 종의 명확한 분류키로 제시한 세포 외각 표면의 그물코 구조(reticulation on the thecal plates)가 Alexandrium sp.(strain WEB-Alex-01)에서는 전혀 없고(Fig. 2-E, F), apical pore plate(Po)와 first apical plate(1')가 완전히 분리된(complete disconnection) A. insuetum 종과는 달리 본 종주는 두 개의 plate가 직접적으로 이어져 있는 유형(direct connection type)의 명확한 형태학적 차이(Fig. 2-F)를 보이고 있다. 따라서 본 종주는 현재까지 종의 동정 및 분류가 명확히 밝혀지지 않은 Alexandrium 신종(new species) 가능성이 높으며, 추가적인 형태학적 특성 분석(SEM, TEM 연구)과 독성 특이성, 계통분류학적 분석 등의 연구가 필요할 것으로 사료된다.
요약 및 결론
(1)국내 연안에서 하계에 출현하는 Alexandrium 속에 속하는 종의 다양성을 파악하기 위해 2013년 하계에 국내 주요 연안지역에서 시료를 채집하여 4종의 Alexandrium 종주를 분리하여 배양체로 확보하였다.
(2)확보한 종주의 형태 및 유전학적 분석 결과, 부산, 태안(가로림만), 광양만에서 채집한 종주는 각각 A. catenella, A. affine, A. fraterculus인 것으로 확인되었는데, 그동안 국내 연안에서는 Alexandrium이 주로 춘계 또는 추계에 진해-마산만 지역에 한정적으로 출현하는 것으로 인식되어 왔으나, 본 연구를 통하여 국내 여러 연안에서 다양한 Alexandrium 종이 하계에도 출현함을 새로이 확인할 수 있었다.
(3)마산만에서 채집한 Alexandrium strain WEB-Alex-01 종주는 국내 연안 최초 출현으로, 차후 이에 대한 신종 분류 연구와 생리, 생태 및 독성에 관한 추가적인 연구가 필요하다.
