

1. 서 론
2. 재료 및 방법
2.1 조사지 개요 및 조사 시기
2.2 환경 요인
2.3 게바다말의 생장 특성
2.4 통계 분석
3. 결 과
3.1 환경 요인
3.2 게바다말 형태적 특성
3.3 게바다말 밀도, 생체량과 생산성
4. 고 찰
1. 서 론
잘피는 지구상의 열대, 아열대, 온대 뿐 만 아니라 일부 냉대 지역의 연안에 출현하며 높은 생산성을 발휘한다(Short et al., 2007). 잘피는 다양한 해양 동물의 직간접적인 먹이원을 공급하고, 잘피 생육지는 이들의 산란장, 은신처와 서식처를 제공하여 생물다양성이 높은 건강한 연안생태계의 필수 구성요소이다(Ruckelshaus and Hays, 1998). 잘피는 무성생식과 유성생식을 통하여 연중 번식하며 개체군을 유지하여 일년생 해조류에서 보이는 끝녹음 현상은 나타나지 않으나, 다양한 환경 요인에 의해 잘피의 생장 형태가 다르게 나타난다(Lee et al., 2005; Lee et al., 2007b).
잘피의 생장에 주로 영향을 미치는 환경요인으로는 수중광량, 수온과 영양염을 들 수 있다. 잘피는 생존과 생장을 위해 원활한 광합성에 필요한 충분한 수중광량이 필요하여 수중광량의 변화에 따라 잘피의 분포와 생장 형태가 달라진다(Dennison, 1987; Lee et al., 2007b). 오랫동안 잘 유지되어오던 잘피 군락지가 갑작스런 적조현상이나, 급격한 부영양화로 플랑크톤이 번성하거나 부착조류가 잘피 잎을 덮어 생존에 필요한 수중광량이 공급되지 못한 상황이 일정 기간 이상 지속되어 소멸되기도 한다(Lee et al., 2007a). 또한 수중광량은 잘피의 생장에 밀접히 연관되어 수중광량이 풍부한 시기에 잘피의 생산성이 증가하고, 수중광량이 감소한 시기에 생산성이 감소한 결과가 보고되기도 한다(Park and Lee, 2009; Park et al., 2016).
수온의 연중 변화는 잘피의 생장이 주기성을 나타내게 작용하며, 이러한 주기성은 잘피가 생육하는 수괴에 따라 잘피 종에 따라 다르게 나타난다(Park et al., 2017). 열대와 아열대지역에 나타나는 Thalassia testudinum이나 Syringodium filiforme은 30°C에서 최대 생장율이 나타난 반면(Gilbert and Clark, 1981), 우리나라를 포함한 북반구 온대지역에 나타나는 거머리말은 15-20°C에서 최대 생장율을 보였다(Lee et al., 2005). 또한, 28°C 이상의 여름 수온이 일정 기간 이상 나타나는 남해연안에 서식하는 거머리말은 고수온의 영향으로 인한 생장 감소가 현저히 나타났지만(Lee et al., 2005), 생육지의 최대 수온이 22°C에 머무르는 동해안의 왕거머리말은 수온이 증가할수록 생장이 활발해졌다(Park et al., 2016). 이처럼 잘피의 광합성과 호흡율에 직접적인 영향을 미치는 수온은 잘피 종과 생육지 환경에 따라 다르게 그 지역에 생육하는 잘피의 생장을 제한한다.
연안에서 높은 일차생산성을 보이는 잘피는 생존하기 위해 풍족한 무기영양염류를 필요로 한다. 대부분의 육상식물은 뿌리로 생존에 필요한 영양염을 흡수하지만, 침수식물인 잘피는 잎과 뿌리로 해수와 퇴적물 공극수의 영양염을 각각 흡수한다(Short, 1987). 생육지의 영양염 환경은 잘피의 생장에 영향을 미쳐 영양염이 풍부한 지역의 잘피가 높은 생산성과 생체량을 나타내는 경우가 빈번하며, 영양염 농도가 낮은 지역에 비료를 시비하여 생장을 촉진시킨 사례도 보고된다(Lee and Dunton, 2000; Lee et al., 2007b). 그러나 해수의 영양염 농도가 높아지면 잘피에 비해 상대적으로 성장이 빠른 플랑크톤이나 조류의 급격한 성장을 초래하여 잘피 잎에 도달하는 수중광량의 감소로 잘피의 생장을 저해시킬 수 있다(Short et al., 1995). 따라서 해수와 퇴적물 공극수의 영양염 환경은 그 곳에 생육하는 잘피의 생장에 직접적 영향을 줄 수 있다.
전 세계 연안에는 60여종의 잘피가 있으며, 대부분은 사니질, 사질 또는 니질로 구성된 연성 저질에 나타난다. 일반적으로 해양 침수식물인 잘피의 지상부 즉 ,잎과 엽초는 해수층에서 수면을 향하여 생장하고, 지하부 즉, 지하경과 뿌리는 부드러운 퇴적물 속에서 수평으로 확장되며 생장한다. 일반적인 잘피와는 달리 말잘피(surfgrass)는 지하경과 뿌리가 퇴적물이 거의 없는 딱딱한 암반에 부착하여 생육하는 독특한 양상을 보인다. 말잘피는 거머리말과(Zosteraceae) 새우말속(Phyllospadix spp.)으로 분류되며 세계적으로 5종이 북태평양연안에 나타난다. 즉, 우리나라, 중국과 일본을 포함하는 북동아시아 연안에는 Asian surfgrass로 불리우는 게바다말(P. japonicus)과 새우말(P. iwatensis)이 출현하고, 미국, 캐나다와 멕시코에 이르는 북미 연안에는 American surfgrass로 불리우는 P. torreyi, P. scouleri와 P. serrulatus가 출현한다(Short et al., 2007). 우리나라 연안의 말잘피 즉, 게바다말과 새우말은 동, 서, 남해안에 모두 출현하지만, 주로 동해안 암반 조하대에 나타나며, 게바다말은 동해안 전역에서 흔히 볼 수 있는데 비해 새우말은 주로 중부 해역에서 제한되어 나타난다(Kim et al., 2018).
최근 갯녹음 현상으로 황폐화가 심각하게 진행되고 있는 우리나라 동해안에서 말잘피의 생태적 중요성이 재조명되면서 이곳의 우점종인 게바다말의 생태적 특성에 관심이 모아지고 있다. 본 연구에서는 동해남부해역의 암반 조하대에 생육하는 게바다말 생육 환경과 생장 특성을 알아보고자 2017년 8월부터 1년 동안 게바다말 군락지의 생육 환경과 게바다말의 형태적 특성, 밀도와 생체량의 변화를 월별 조사하였다. 또한, 동해안에서 조사된 새우말속 잘피의 생태적 특성을 비교하여 이 지역 게바다말의 생장 특성을 밝히고자 한다.
2. 재료 및 방법
2.1 조사지 개요 및 조사 시기
본 연구는 우리나라의 동해남부해역에 속하는 경북 포항시 구룡포 연안(36° 00′ 25″ N, 129° 34′ 43″ E)의 게바다말 군락지에서 이루어졌다. 이 지역은 반일주조로 조차는 평균 30 cm이며, 수심 2-5 m의 암반에 단일종의 말잘피, 게바다말 군락이 넓게 형성되어 있다(Fig. 1). 게바다말 군락지의 생육 환경 즉, 수중광량, 수온과 영양염 농도, 그리고 게바다말의 생태적 특성 즉, 밀도, 생체량, 잎 생산성과 형태적 특성의 조사는 2017년 8월부터 2018년 7월까지 진행되었다.
2.2 환경 요인
게바다말 군락지의 수중광량과 수온을 조사하기 위해 게바다말의 잎이 위치한 수심에 HOBO data logger (Onset Computer Corp., Bourne, MA, UAS) 센서를 설치하여 매 15분마다 측정하였다. 사용된 모든 센서는 LI-1400 Data logger와 LI-193 SA 구형 광량센서(Li-Cor, Inc, Lincoln, NE, USA)를 이용하여 동시 측정한 광량값과 회귀분석을 통해 조도(lumen ft-2) 측정치를 수중광량으로 변환하였다(Lee et al., 2005; Park and Lee, 2009). 조사 장소에 시설된 센서는 월 2회 교체되었으며, 그 때마다 측정 자료를 수집하여 수중광량과 수온의 월 평균값으로 나타내었다. 게바다말 군락지의 영양염 농도를 분석하기 위해 월 1회 게바다말 군락지에서 해수 120 mL을 채수하여(n=4) 냉장 보관 후 실험실로 옮긴 후 박막여과지(cellulose membrane filters, Milipore)로 여과 후 암모늄염(NH4+), 질산염+아질산염(NO3-+NO2-)과 인산염(PO43-)을 흡광광도법을 이용하여 측정하였다(Parsons et al., 1984).
2.3 게바다말의 생장 특성
게바다말의 형태적 특성, 밀도, 생체량과 생산성의 변화를 매월 조사하였으며, 채취한 모든 시료는 얼음이 담긴 아이스박스에 보관하여 실험실로 운반하였다. 형태적 특성의 월 변화를 보기 위해 조사 시기마다 8-10개의 성숙한 게바다말 시료를 채취하여 잎의 수, 잎 폭과 두께, 엽초 길이와 폭 및 개체 길이를 측정하였다. 잎의 수는 각 개체의 엽초 위로 출현한 모든 잎 수를 헤아렸고, 잎 폭(mm)과 두께(mm)는 3번째 잎 중간 부분의 폭과 두께를 각각 측정하였다. 엽초 길이(cm)는 생장점부터 엽초 상단 경계까지의 길이, 엽초 폭(mm)은 엽초의 가장 넓은 부분의 폭을 측정하였다. 개체 길이(cm)는 생장점부터 가장 긴 잎 상단까지의 길이를 측정하였다.
밀도의 변화를 측정하기 위해 매월 임의로 던진 방형구(50×50 cm, n=4)내의 모든 개체를 영양지와 육수화서로 구분하여 단위면적당 개체수로 나타내었다. 생체량의 변화를 보기 위해 매월 임의로 놓은 방형구(10×10 cm, n=4)내 모든 게바다말을 채취하여 흐르는 물에 세척하면서 이물질을 제거하여 지상부(잎과 엽초)와 지하부(지하경과 뿌리)로 구분한 후 60°C에서 48시간 이상 건조한 후 단위면적당 건중량(g DW m-2)으로 나타내었다. 잎 생산성의 변화는 잎 표시법(leaf marking method)을 이용하여 측정하였다(Lee et al., 2005; Park and Lee, 2009). 즉, 매월 임의로 선정된 게바다말 성체의 엽초 부분에 날카로운 주사바늘로 구멍을 뚫은 후(n=6-10) 약 1개월 후 회수하여 기존 부분(구멍 뚫기 전)과 새로 자란 부분(구멍 뚫은 후)으로 구분 후 60°C에서 항량이 될 때까지 건조하였다. 개체당 잎 생산성(mg DW sht-1 d-1)은 새로 자라난 잎 중량을 실험일수로 나누어 구하였으며, 여기에 매월 측정된 생육 밀도를 곱하여 단위면적당 잎 생산성(g DW m-2 d-1)을 구하였다.
2.4 통계 분석
모든 측정치는 평균값(mean)과 표준오차(SE)로 나타내었다. 통계분석은 SPSS10.1 버전(SPSS, Inc., Chicago, IL, USA)을 이용하였으며, 모든 자료는 normality와 homogeneity of variance를 검정한 후 one-way ANOVA를 이용하여 조사 시기에 따른 차이의 유의성을 검사하였다. 환경 요인과 말잘피 측정치의 상관성은 피어슨 상관관계 분석을 이용하여 평가하였다.
3. 결 과
3.1 환경 요인
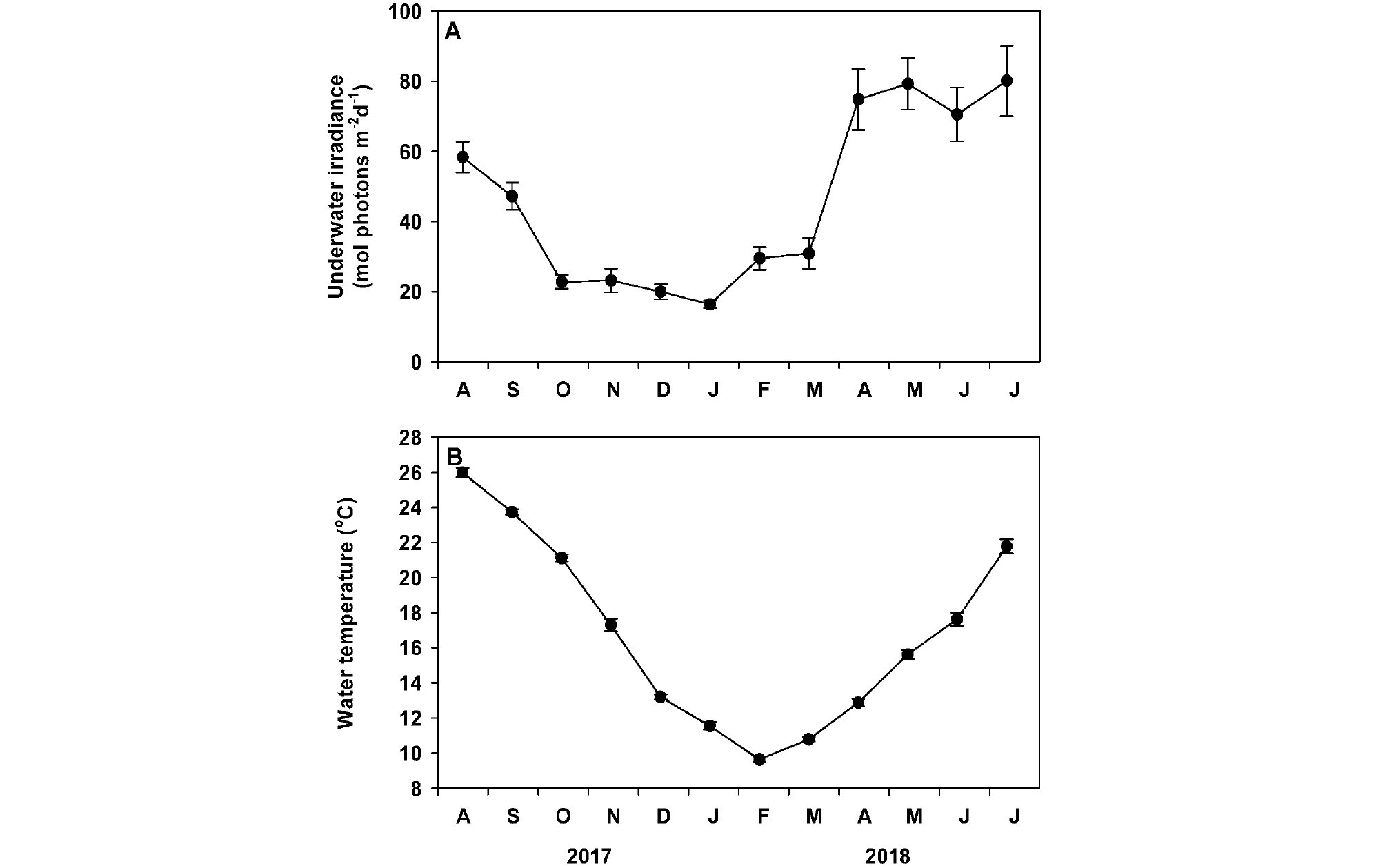
게바다말 군락지에서 측정된 수중광량은 조사 시기에 따라 유의한 차이(p <0.001)가 나타났으며, 봄과 여름에 증가하고 가을과 겨울에 감소하는 뚜렷한 계절 경향을 나타내었다(Fig. 2A). 조사 기간 중 평균 수중광량은 46.1±7.3 mol photons m-2 d-1로 나타났다. 월평균 수중광량은 2018년 1월에 가장 낮았으며(16.4±1.1 mol photons m-2 d-1), 2018년 7월에 가장 높았다(80.1±10.0 mol photons m-2 d-1, Fig. 2A). 수온도 가을과 겨울에 감소하고 봄과 여름에 증가하는 뚜렷한 계절 경향이 나타났다. 게바다말 군락지의 조사 기간 중 평균 수온은 16.8±1.6°C로 기록되었다. 월 평균 수온은 2018년 2월에 최소값(9.6±0.2°C)을 보였으며, 2017년 8월에 최대값(26.0±0.3°C)을 나타내었다(Fig. 2B).
게바다말 군락지에서 측정된 해수의 영양염 농도는 조사 시기에 따라 유의한 차이(p <0.001)가 나타났다(Table 1, Fig. 3). 조사 기간 중 수층의 평균 암모늄염 농도는 2.4±0.4 μM로 나타났다(Fig. 3A). 평균 질산염+아질산염과 인산염의 농도는 각각 1.3±0.2 μM과 0.4±0.0 μM로 나타났다(Figs. 3B, 3C).
Table 1. Results of a one-way ANOVA among sampling times on water column nutrients, shoot density, productivity, biomass, and shoot morphology

3.2 게바다말 형태적 특성
게바다말의 형태(즉, 잎 수, 잎 폭과 두께, 엽초 길이와 폭, 개체 길이)는 조사 시기에 따라 유의한 차이(p <0.001)가 나타났다(Table 1, Fig. 4). 잎 수는 평균 4.8±0.1개로 2017년 10월에 가장 작았고(3.9±0.2개), 2018년 3월, 5월과 6월에 많았다(5.1±0.1개, Fig. 4A). 잎 폭은 평균 2.3±0.1 mm로, 2017년 9월에 최소값(1.9±0.1 mm)을 나타낸 후 증가하여 2018년 2월에 최대값(2.6±0.1 mm)이 나타났다(Fig. 4B). 잎 두께는 조사 기간 중 평균 0.33±0.01 mm로 2018년 3월에 가장 얇았고(0.27±0.01 mm), 2017년 10월에 가장 두꺼웠다(0.39±0.01 mm, Fig. 4C).
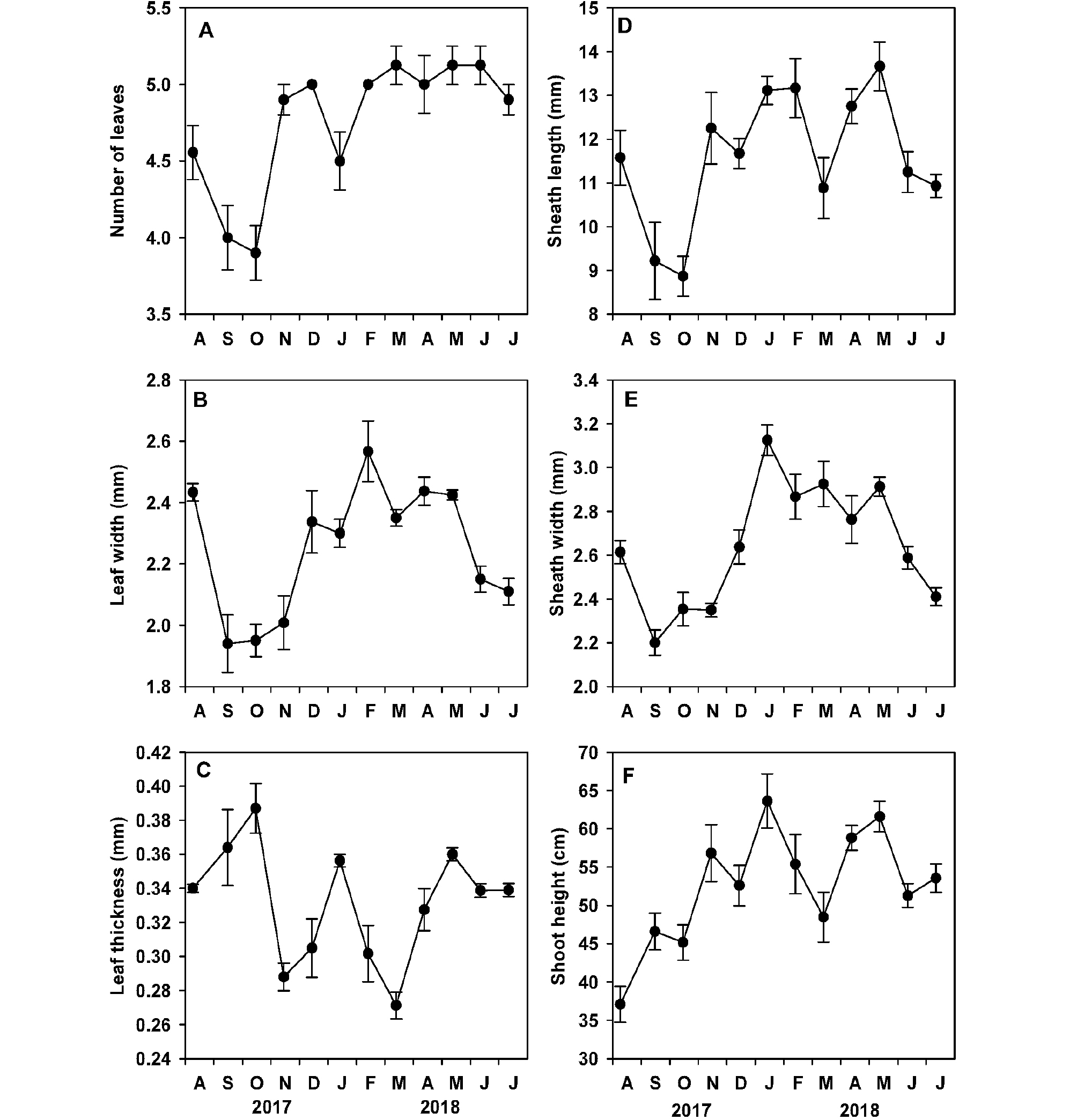
엽초 길이는 평균 11.6±0.4 cm로, 2017년 10월에 가장 짧았고(8.9±0.5 cm), 2018년 5월에 가장 길었다(13.7+0.6 cm, Fig. 4D). 엽초 폭은 2017년 9월에 최소값(2.2±0.1 mm)과 2018년 1월에 최대값(3.1±0.1 mm)을 보였으며, 조사 기간 중 평균 2.6±0.1 mm로 나타났다(Fig. 4E). 개체 길이는 2017년 8월 가장 짧았다가(37.1±2.4 cm) 서서히 증가하여 2018년 1월 최대값(63.6±3.5 cm)을 나타내었으며, 조사 기간 중 평균 52.6±2.2 cm이었다(Fig. 4F). 개체 길이는 수중광량과 유의한 양의 상관관계가 나타나지 않았고(r=0.023, p=0.944), 수온과 약한 음의 상관관계(r=-0.670, p <0.05)를 나타내었다.
3.3 게바다말 밀도, 생체량과 생산성
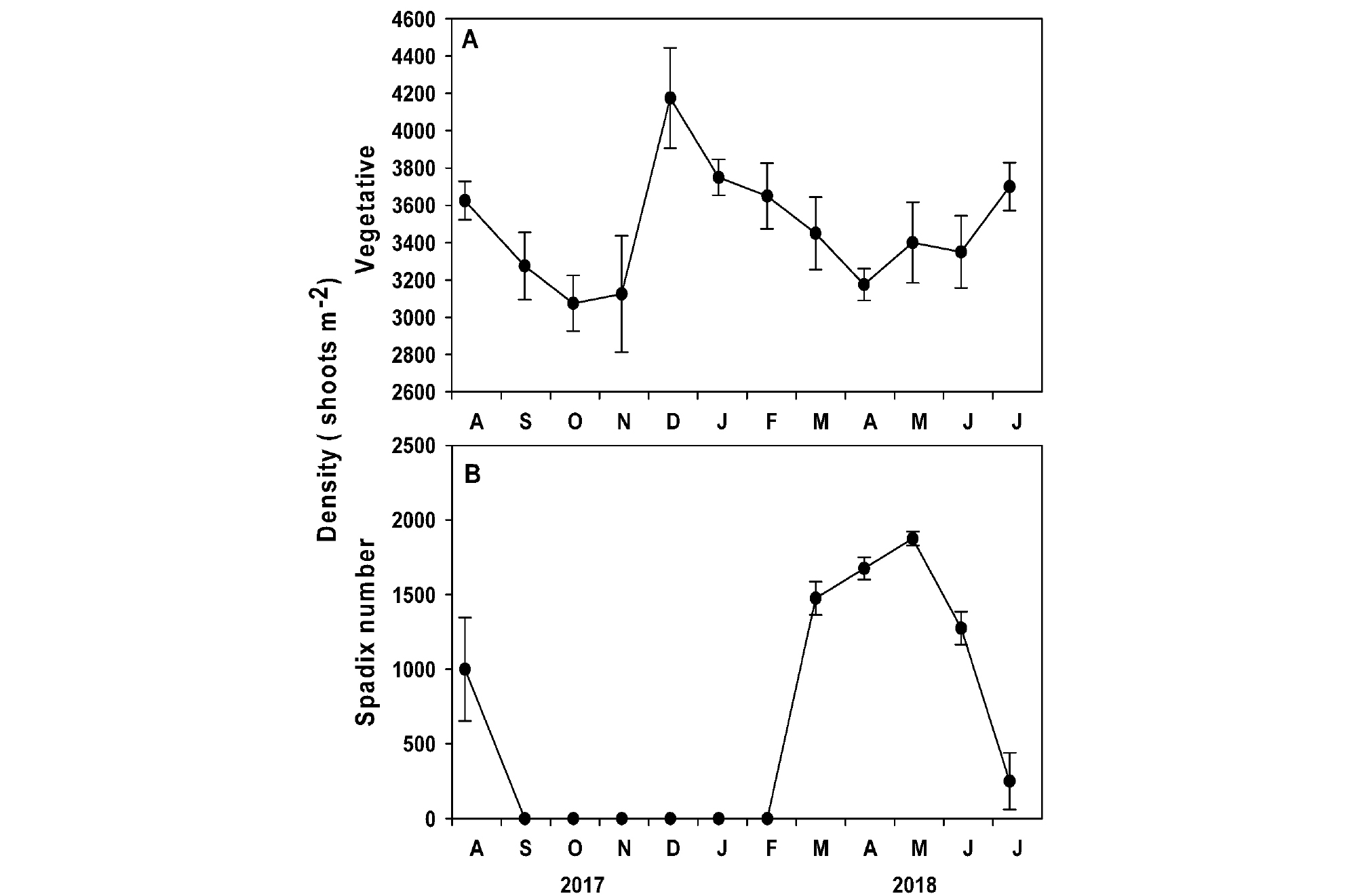
게바다말 영양지와 육수화서 밀도는 조사 시기에 따라 유의한 차이(영양지: p <0.05, 육수화서: p <0.001)가 발생하였다(Table 1, Fig. 5). 게바다말 영양지의 평균 밀도는 3479.2±91.4 shoot m-2로 2017년 10월에 가장 낮았고(3075.0±149.3 shoot m-2), 2017년 12월에 가장 높았다(4175.0±268.9 shoot m-2,Fig. 5A). 육수화서는 2018년 3월부터 관찰되기 시작하여 5월에 최대밀도(1875.0±47.9 shoot m-2)를 나타낸 후 8월 이후 소멸되었으며, 육수화서 밀도는 수중광량과 유의한 양의 상관관계(r=0.661, p <0.05)가 나타났다(Fig. 5B).
게바다말 지상부와 지하부의 단위면적당 생체량은 조사 시기별 유의한 차이(p <0.001)가 나타났으며, 가을에 낮고 봄에 높은 경향을 보였다(Table 1, Fig. 6). 지상부 생체량은 2017년 11월에 최소값(654.5±54.8 g DW m-2)과 2018년 5월에 최대값(1583.7±127.5 g DW m-2)이 나타났으며, 조사 기간 평균 1158.1±81.5 g DW m-2로 단위면적당 생체량의 약 68.2%를 차지하였다(Fig. 6A). 지하부의 생체량은 529.6+25.2 g DW m-2로 단위면적당 생체량의 약 31.8%를 차지하였으며, 2017년 11월에 최소값(407.5±34.7 g DW m-2)과 2018년 6월에 최대값(647.9±34.0 g DW m-2)이 나타났다(Fig. 6B). 게바다말의 단위면적당 평균 생체량은 1687.8±103.2 g DW m-2이었으며, 2017년 11월에 최소값(1062.0±87.0 g DW m-2)과 2018년 5월에 최대값(2195.8±133.2 g DW m-2)이 나타났다(Fig. 6C).
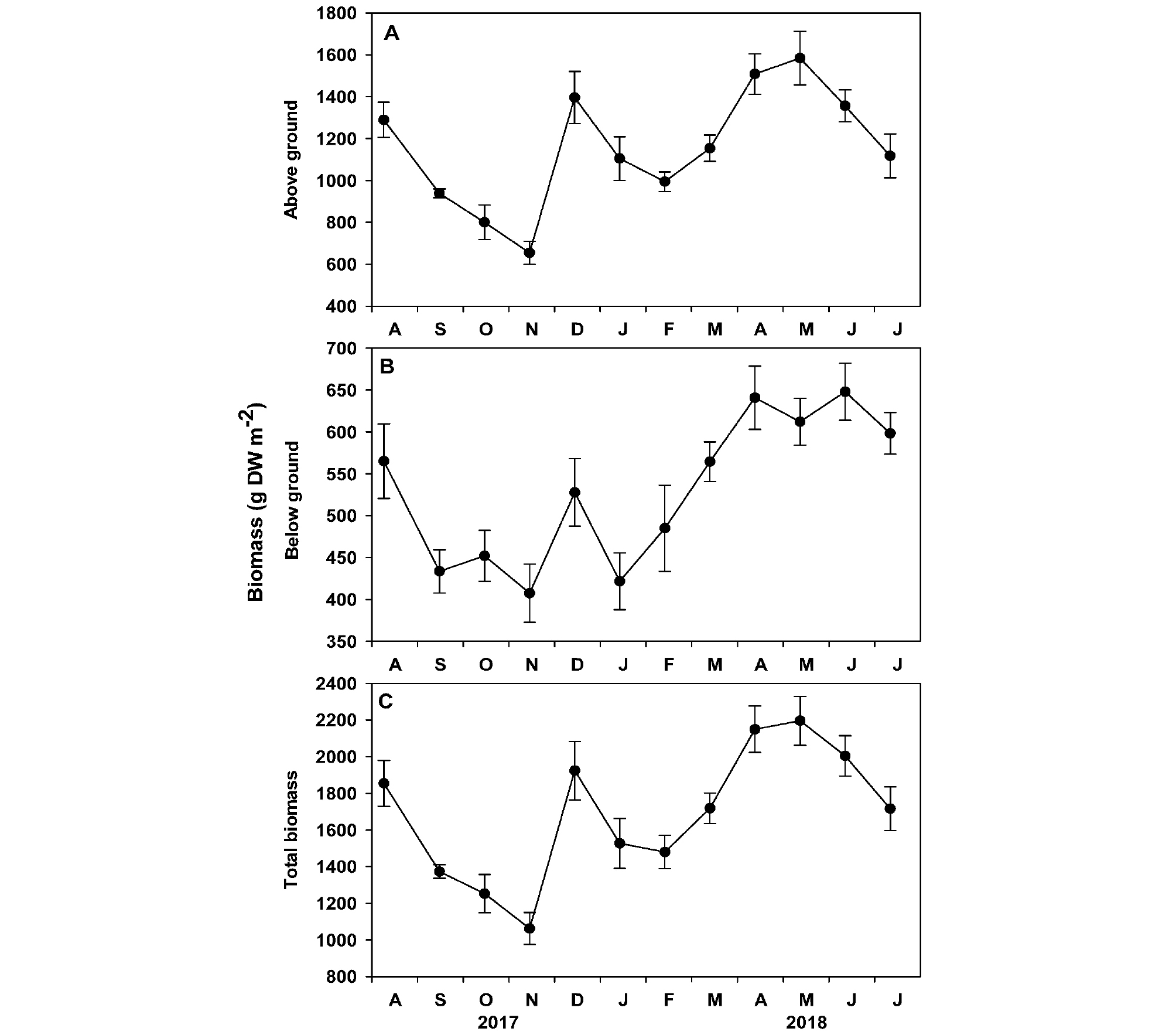
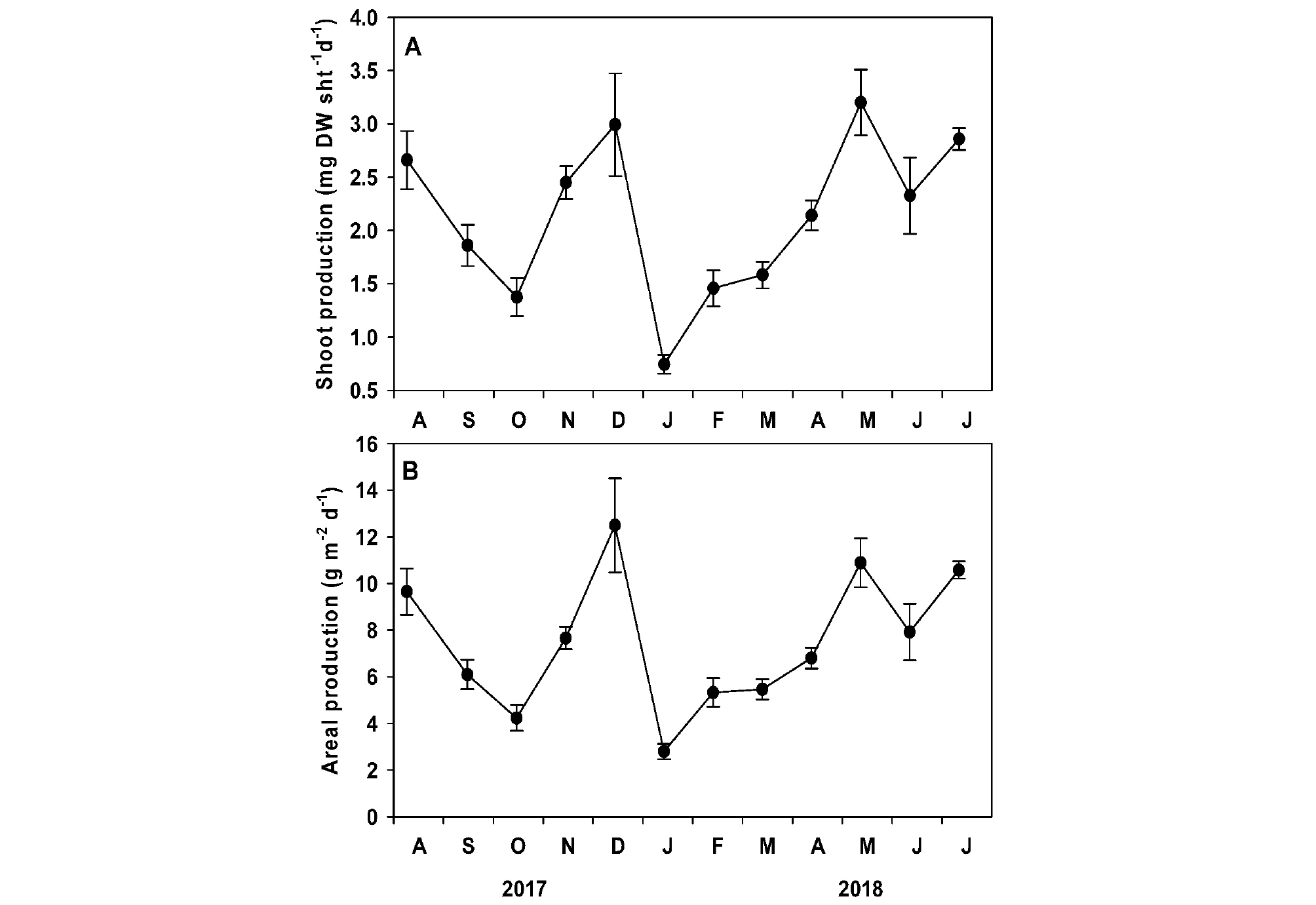
게바다말의 개체당 잎 생산성과 단위면적당 잎 생산성은 조사 시기에 따라 유의한 차이(p <0.001)가 나타났다(Table 1, Fig. 7). 게바다말의 평균 개체당 잎 생산성은 2.1±0.2 mg DW sht-1 d-1로 초겨울인 2017년 12월 이후 1달 동안 급격히 낮아진 후 서서히 증가하여 2018년 5월 최대값(3.2±0.3 mg DW sht-1 d-1)이 나타났다(Fig. 7A). 게바다말의 개체당 잎 생산성은 수중광량(r=0.566, p= 0.055), 수온(r=0.349, p= 0.267), 해수의 영양염 농도(암모늄염: r=-0.490, p= 0.880, 질산+아질산염: r=-0.268, p= 0.399, 인산염: r=-0.346, p= 0.271)와 유의한 상관관계가 나타나지 않았다. 단위면적당 잎 생산성도 개체당 잎 생산성과 유사한 경향이 나타났으며, 조사 기간 평균 7.5±0.8 g DW m-2 d-1로, 2018년 1월에 최소값(2.8±0.3 g DW m-2 d-1)과 2017년 12월에 최대값(12.5±2.0 g DW m-2 d-1)을 나타났다(Fig. 7B). 조사 지역 게바다말의 개체당 잎 생산성은 수중광량(r=0.468, p= 0.125), 수온(r=0.256, p= 0.422), 해수의 영양염 농도(암모늄염: r=-0.179, p= 0.577, 질산+아질산염: r=-0.200, p= 0.534, 인산염: r=-0.451, p= 0.141)와 유의한 상관관계가 나타나지 않았다.
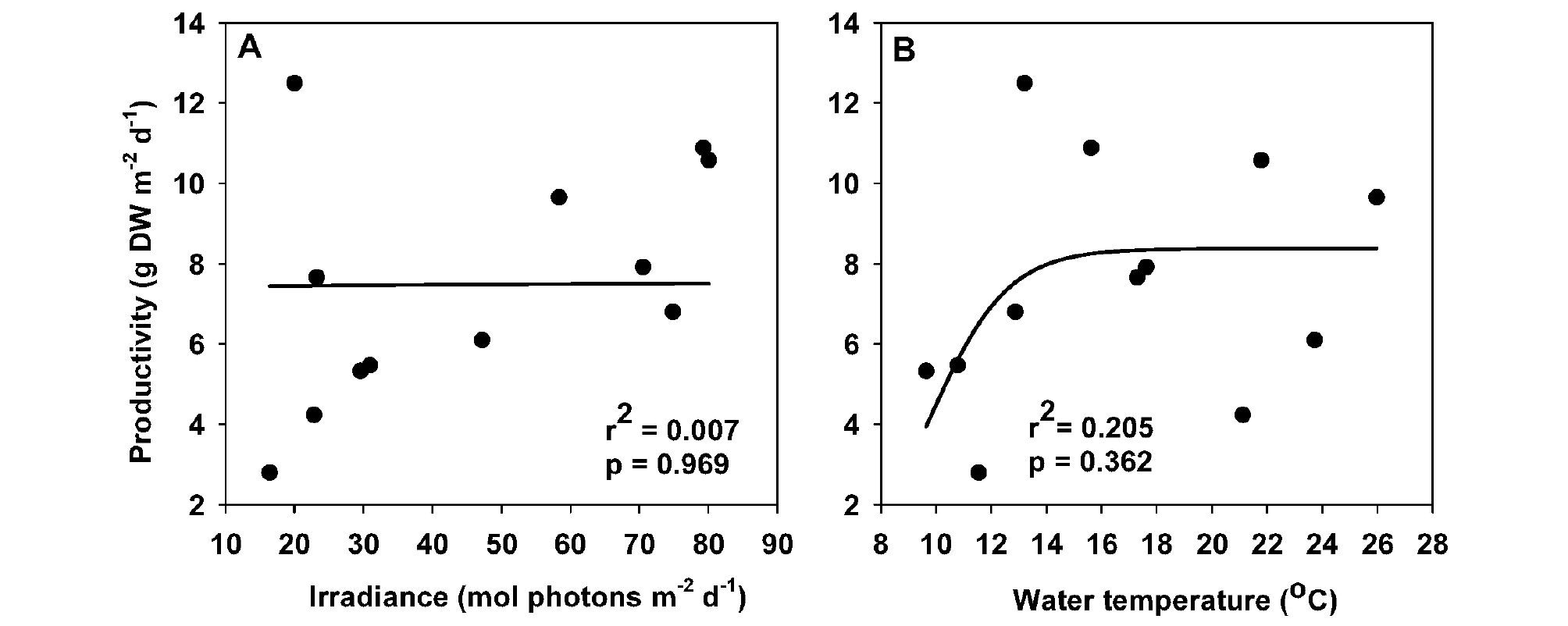
게바다말의 단위면적당 잎 생산성을 각각 수중광량 및 수온과 비선형 회귀분석을 실시하면, 이 지역 게바다말의 생산성은 수중광량(r2=0.007, p =0.969; Fig. 8A)이나 수온(r2=0.205, p =0.362; Fig. 8B)과는 유의한 상관성이 나타나지 않았다.
4. 고 찰
수중광량은 잘피의 생존, 생장과 분포를 결정하는 주요 제한 요소이다. 침수식물인 잘피는 종에 따라 동종이라도 생육 환경에 따라 요구되는 수중광량의 차이를 보인다. 타원형의 잎을 가진 해호말은 피침 모양의 잎을 가진 잘피 종보다 수중 빛을 포획하는데 유리하여 깊은 수심에서도 생존이 가능하다(Kuo and den Hartog, 2006). 또한, 피침 모양의 잎을 가진 잘피 종에서는 넓은 잎 폭을 가진 종이 좁은 잎 폭을 가진 종보다 수중 빛의 포획에 유리하였다(Smith et al., 1997; Park et al., 2016). 남해안 거머리말 군락지(11.7-28.0 mol photons m-2 d-1)의 수중광량이 동해안 새우말 군락지(39.3 mol photons m-2 d-1, Table 2)보다 현저히 낮음에도 불구하고, 남해안 거머리말의 생장은 수중광량에 의해 제한받지 않았으나(Lee et al., 2005), 동해안 새우말의 생장은 수중광량의 유의한 영향을 받았다(Kim et al., 2018). 즉, 거머리말속과 새우말속은 유사한 피침모양의 잎을 지니고 있으며, 새우말속에 비해 거머리말 속은 넓은 잎 폭을 보여 수중 빛의 포획에 유리하게 작용할 수 있었을 것이다. 또한, 남해안 거머리말 군락지보다 비교적 높은 수중광량을 보였던 부산 연안의 게바다말 군락(24.7 mol photons m-2 d-1, Table 2)에서도 잎 생산성과 생육지의 수중광량은 유의한 양의 상관관계를 보였다(Park and Lee, 2009). 따라서 새우말속은 거머리말속보다 생장에 필요한 최소 광요구량이 높을 것으로 추측할 수 있다. 그러나 조사장소의 게바다말 군락의 개체당 잎 생산성과 면적당 잎 생산성은 수중광량과 유의한 관계가 나타나지 않았다(Fig. 8A). 이는 이 지역 게바다말은 조사 기간 동안 충분한 수중광량을 공급받아 게바다말의 생장이 광에 의해 결정적으로 제한되지 않았음을 의미한다.
Table 2. Mean values (ranges) of physical and chemical parameters, biological measurements for surfgrass, Phyllospadix japonicus and P. iwatensis on the eastern coast of Korea
| Parameters | This study | P. japonicus | P. iwatensis |
| Underwater irradiance | 46.1 (16.4-80.1) | 24.7 (7.8-48.1) | 38.7 (10.9-78.3) |
| (mol photons m-2 d-1) | |||
| Water temperature (°C) | 16.8 (9.6-26.0) | 18.3 (11.5-26.4) | 14.4 (5.1-23.9) |
| Water column nutrients(μM) | |||
| NH4+ | 2.4 (0.9-5.3) | 4.1 (0.9-16.3) | - |
| NO3-+NO2- | 1.3 (0.3-2.1) | 3.4 (0.3-10.8) | - |
| PO43- | 0.4 (0.2-0.6) | 0.8 (0.3-2.8) | - |
| Shoot density (shoots m-2) | 4108.3 (3075.0-5275.0) | 937.8 (676.3-1290.0) | 2416.7 (1900.0-2900.0) |
| Spadix ratio (%) | 15.3 (0.0-35.5) | 6.3 (0.0-25.0) | 4.4 (0.0-16.4) |
| Productivity | |||
| Leaf productivity | 2.1 (0.7-3.2) | 4.5 (1.7-7.7) | 2.7 (0.9-6.6) |
| (mg DW shoot-1 d-1) | |||
| Areal productivity | 7.5 (2.8-12.5) | 4.0 (1.3-9.3) | 6.3 (2.4-16.4) |
| (g DW m-2 d-1) | |||
| Biomass (g DW m-2) | |||
| Leaf | 1158.1 (654.5-1583.7) | 487.8 (177.6-1163.0) | 735.6 (616.1-1001.4) |
| Rhizome + root | 529.6 (407.5-647.9) | 211.5 (98.7-387.2) | 631.7 (514.9-844.2) |
| Total biomass | 1687.8 (1062.0-2195.8) | 699.6 (305.6-1551.0) | 1367.4 (1131.0-1691.3) |
| Shoot morphology | |||
| Number of leaves | 4.8 (3.9-5.1) | 4.6 (3.9-5.2) | 5.1 (4.6-5.9) |
| Leaf width | 2.3 (1.9-2.6) | 2.4 (1.9-2.9) | 3.0 (2.8-3.3) |
| Leaf thickness | 0.33 (0.27-0.39) | - | 0.30 (0.25-0.35) |
| Shoot height | 52.6 (37.1-63.6) | 70.0 (34.5-98.3) | 50.2 (34.1-69.3) |
| Reference | Park and Lee, 2009 | Kim et al., 2016 |
수온의 계절 변화는 잘피의 생장이 계절적 주기를 보이게 작용한다. 조사 장소 게바다말의 형태적 특성, 밀도, 생산성과 생체량은 여름 이후 감소하다가 월 평균 수온이 13.2°C인 2017년 12월에 급격히 높은 값을 보였다가 감소한 후 봄에 다시 증가하였다. 생육지 수온이 높은 시기에 잘피 생장이 감소하고, 생육지 수온이 최저값을 나타낸 후 증가하면서 잘피 생장이 활발한 생장 형태는 온대 수괴에 서식하는 잘피의 일반적인 생장 특성이다. 온대 잘피의 생장에 적합한 수온은 수온이 증가하는 시기의 11.5-26°C로 보고되며, 수온이 더 증가하게 되면 호흡율의 증가로 잘피 생장이 저하가 나타난다(Lee et al., 2007b). 우리나라 남해안의 만에 주로 서식하는 거머리말의 생장은 수온이 15-20°C를 나타내는 초봄부터 초여름까지 가장 활발하며, 수온이 더 증가하면 생장이 감소하기 시작하고 고수온에서는 광합성률의 증가율보다 호흡률의 증가율이 더 커져 고수온 장애가 나타난다(Lee et al., 2005). 생육지 평균 수온이 본 조사 장소보다 약 1.5°C 높은 부산 연안의 게바다말 군락의 잎 생산성은 약 14°C에서 최대값을 보인 후 수온의 증가와 함께 감소하는 상관관계가 나타났다(Park and Lee, 2009). 그러나 생육지 평균 수온이 본 조사 장소보다 약 2.4°C 낮은 강원도 고성 새우말 군락의 잎 생산성은 12-13°C에서 최대값을 보였고 수온의 증가로 유의한 생산성의 감소는 나타나지 않았다(Kim et al., 2018). 본 조사 장소의 게바다말의 잎 생산성은 13.2°C에서 최대값이 나타났으나, 수온의 증가로 인한 유의한 생산성의 감소는 나타나지 않아(Fig. 8B) 이 지역 게바다말의 생장은 수온에 의해 결정적으로 제한받지 않았다고 볼 수 있다. 생육지 수온이 25°C를 초과하지 않는 조하대의 새우말 군락지에서는 고수온으로 인한 스트레스가 발생하지 않았다(Yabe et al., 1996). 최대 생장을 보이는 수온이 새우말보다 조금 높게 나타나는 게바다말은 이 지역 생육지 수온이 26°C를 초과하지 않아 고수온으로 인한 스트레스가 나타나지 않은 것으로 추측된다.
연약지반에 서식하는 잘피는 잎과 뿌리로 해수와 퇴적물 공극수의 영양염을 흡수하여 생장하므로, 서식지 수층과 퇴적물의 영양염 농도는 잘피의 생장을 제한하기도 한다(Lee and Dunton, 2000). 그에 비해 퇴적물이 거의 없는 암반에 부착하여 생장하는 새우말속 잘피는 잎으로 수층의 영양염을 흡수하여 생장한다(Turner, 1983; Terrados and Williams, 1998). 본 조사의 게바다말의 잎 생산성은 군락지 해수의 영양염 농도와 유의한 상관관계가 나타나지 않아, 이 지역 게바다말 군락의 생장이 영양염에 의해 제한되지는 않았다고 볼 수 있다. 이러한 해수 영양염 농도와 게바다말의 생장과의 상관관계는 부산 연안의 게바다말 군락지에서도 유사하게 나타났다(Park and Lee, 2009).
말잘피는 높은 육수화서 비율을 보이는 것으로 알려져 있으며, 종과 생육지 환경에 따라 그 비율은 다르게 나타난다(Williams, 1995; Ramírez-García et al., 2002). 육상식물에 비해 빛의 공급이 불리한 침수식물인 잘피는 화지와 생식 기관의 형성에 많은 에너지를 필요로 한다(Kaldy, 1999). 연성 저질에 서식하는 잘피는 잎 뿐 만아니라 뿌리로도 영양염을 흡수하여 지하경에 탄소를 저장하며, 에너지가 필요한 화지 형성 시기에 지하경에 저장된 탄소를 이용한다(Marbà et al., 2002). 그러나 파도가 심한 해역의 암반에 부착하여 살아가는 말잘피의 지하경과 뿌리는 탄소의 저장고 역할보다는 센 파도와 조류로부터 탈락되지 않기 위해 암반에 부착하는 기능을 수행한다(Hasegawa et al., 2005). 따라서 말잘피는 육수화서와 생식 기관이 형성되는데 필요한 에너지의 이용에 지하경의 저장 탄소를 활용하는데 제한적일 수 있다. 본 조사 장소 게바다말의 평균 육수화서 비율은 15.3%로 부산 연안 게바다말(6.3%)과 고성 새우말(4.4%)보다 월등히 높았다(Table 2). 조사 장소 게바다말의 형태적 특성, 밀도, 생산성과 생체량이 초겨울인 12월에 최대를 보인 후 급격히 감소하는 특이한 현상이 뚜렷이 나타난 것은 다량의 육수화서를 발생시키기에 필요한 에너지 공급과 연관될 수 있을 것이다.
결론적으로 조사 장소의 게바다말은 가을에 생장이 감소하고 봄에 증가하여 동해안에 생육하는 새우말속 잘피와 유사한 생장 특성이 나타났다. 그러나, 조사 장소 게바다말 군락의 잎 생산성이 최대로 나타난 수온은 13-14°C로 부산 연안 게바다말 군락(약 14°C)보다 조금 낮았고, 고성군 새우말 군락(12-13°C)보다는 조금 높았다. 부산 연안 게바다말의 생산성은 생육지의 수중광량과 수온에 유의한 상관관계가 나타났고, 고성 새우말의 생산성은 생육지의 수중광량과 유의한 상관관계를 보였으나, 조사장소 게바다말의 생산성은 생육지의 수중광량, 수온이나 해수의 영양염 농도에 결정적으로 제한받지 않았다. 이는 조사 기간 중 이 곳 게바다말 군락지는 게바다말 생장에 필요한 충분한 빛이 공급되고, 적절한 수온 범위와 영양염 농도가 유지되었음을 알 수 있다.
