

1. 서 론
반원형 형태의 Amphora Ehrenberg ex Kützing 속은 오랫동안 여러 학자에 의해 단일 계통군이 아닌 다계통군이라 주장되었다(Smith, 1873; Cleve, 1895; Mereschkowsky, 1903) Cleve(1895)는 Amphora 속내 종들이 형태 차이가 있는 여러 분류군으로 이루어진 것으로 판단하여, 각(frustule)과 둘레띠(girdle)의 형태 특징을 기반으로 Archiamphora, Amblyamphora, Amphora, Calamphora, Cymbamphora, Diplamphora, Halamphora, Oxyamphora, Psammamphora 등 9개 아속으로 구분하였다. Mereschkowsky(1903)는 Amphora 속의 엽록체 형태를 9개 유형으로 나누고 4개 층으로 된 단일 엽록체가 배부에 위치한 제1 유형의 Clevamphora Mereschkowsky와 배부의 2개 엽록체와 등부의 2개 엽록체가 피레노이드로 연결되어 쌍을 이룬 제8 유형의 Tetramphora Mereschkowsky을 신속으로 제안하였다. Mereschkowsky의 제안은 오랫동안 간과 되어 왔고, 대부분의 연구자들은 Cleve(1895)가 제안한 9개의 아속 체계를 채택해왔다. 이후 다계통인 Amphora의 계통분류학적 개정이 시도되면서(예, Williams and Reid, 2006; Stepanek and Kociolek, 2014), Tetramphora 속 역시 연구자들의 관심을 받게 되었다(Stepanek and Kociolek, 2016). Stepanek and Kociolek(2016)은 비대칭적인 반원형 세포, 세포 양쪽에 아치 형태로 분포하는 등줄(raphe), 중앙부의 규산질 돌기 그리고, Mereschkowsky의 제8 유형 엽록체 구조 등에 따라 Tetramphora 속의 특성을 수정하였다.
Tetramphora는 현재 12종이 알려졌고(Guiry and Guiry, 2024) 국내에는 Amphora로 보고된 T. rhombica, T. decussata (심, 1994), T. ostrearia (Park and Koh, 2012) 3종과 Tetramphora로 보고된 T. intermedia (Park et al., 2017) 1종을 포함하여 모두 4종이 분포하는 것으로 알려졌다. 본 연구에서는 2021년 5월에 포항에서 발견한 T. sulcata를 보고한다.
2. 재료 및 방법
2.1 조사 및 단일 세포 분리 배양
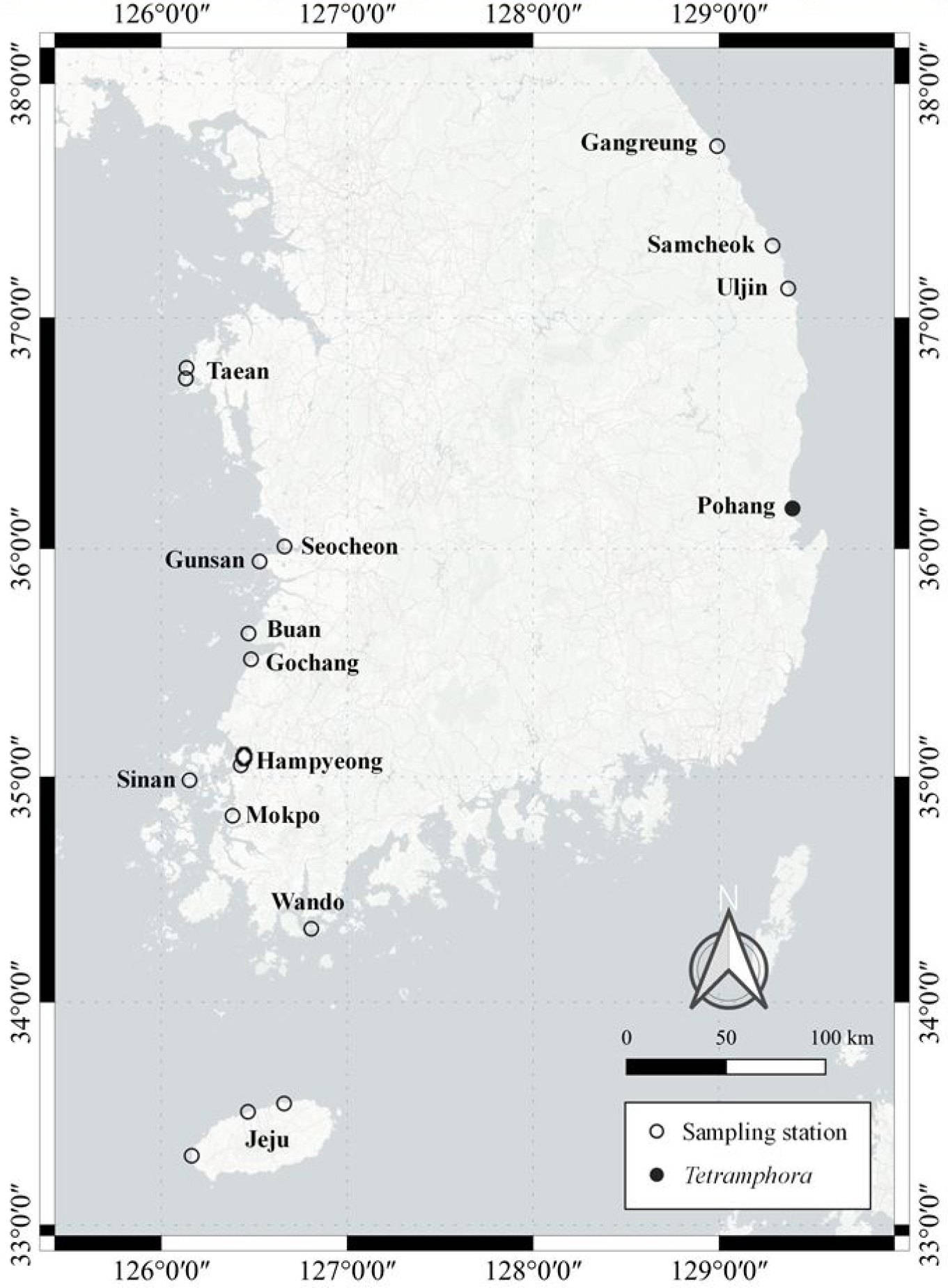
연안 저서규조류 연구를 위해 전국 18개 정점에서 시료를 채집하였으며, 본 연구에서 보고하는 종은 2021년 5월 경상북도 포항시 북구 청하면 청진2리(36°10′30″N, 129°23′43″E)에서 확보한 시료에서 발견되었다(Fig. 1). 간조 시 노출된 해빈에서 자갈을 채집하여 100 mL 폴리에틸렌(PE) 재질의 채수병에 밀봉 보관하였고, 모래 시료는 표층으로부터 1 cm 깊이에서 Petri dish를 이용해 채집한 뒤 동일한 방식으로 밀봉하여 실험실로 운반하였다.
운반된 자갈 및 모래 시료는 도립현미경(CK40, Olympus, Japan) 하에서 파스퇴르 피펫을 사용하여 Tetramphora의 단일 세포를 분리하였다. 분리된 세포는 f/2-Si 배지가 담긴 24-well 세포배양용 culture plate에 접종하였으며, 일정 수준으로 생장한 후 125 mL PC bottle (Nalgene, USA)에 100 mL의 f/2-Si 배지를 담아 옮겨 배양을 진행하였다. 배양 조건은 광도 100 μmol photons m-2 s-1, 광주기 16:8 (명:암), 온도 18℃, 염분 32 psu로 설정하였다
2.2 형태 관찰
규조류의 형태는 광학현미경(Axioplan, Carl Zeiss, Germany) 하에서 400–1000배 배율로 미분간섭대조(Differential Interference Contrast, DIC) 및 명시야(Bright-Field, BF) 방식으로 관찰하였다. 세포 내 유기물을 제거하고 규산질 껍질의 미세 구조를 관찰하기 위한 전처리 과정은 Hasle and Fryxell(1970)의 방법을 따랐다. 전처리된 시료는 일부를 피펫으로 채취하여 커버글라스에 옮긴 뒤 건조하고, 슬라이드글라스 위에 Naphrax (Brunel Microscopes Ltd., Chippenham, UK)를 사용하여 봉입하여 영구 프레파라트 표본으로 제작하였다. 해당 표본 내 Tetramphora 시료를 대상으로 세포의 크기, 상각(epivalve) 및 하각(hypovalve) 끝의 모양, 세포의 전체 형태를 관찰하였다. 엽록체는 살아있는 배양 시료를 커버글라스로 옮긴 후 바로 촬영하여 관찰하였다.
전자현미경 관찰을 위해서는 배양 시료를 3% Lugol 용액으로 고정하였다. 전처리 시료는 해수와 증류수를 서로 다른 비율로 혼합하여 염분을 제거하였으며, 에탄올을 사용하여 수분을 제거하였다. 완전한 에탄올 치환을 위해 100% 에탄올에서 1시간 동안 탈수 과정을 수행하였다. 에탄올로 치환된 시료는 Petri dish에 담아 밀봉하여 보관하였으며, 전자현미경 관찰 직전에는 Ion Sputter (E-1045, Hitachi, Japan)를 이용하여 15 mA 전류 조건에서 40초간 백금(Pt) 코팅을 실시하여 전도성을 부여하였다. 이후, 가속 전압 3 kV 조건에서 전자현미경(S-4800+EDS, Horiba: EX-250, Hitachi, Japan)을 이용하여 관찰을 수행하였다.
2.3 유전자 분석
배양주 50 mL를 3,000 rpm에서 20분간 원심분리한 후, DNeasy Plant Mini Kit (Qiagen, Valencia, CA, USA)를 이용하여 DNA를 추출하였다. rbcL 유전자 분석을 위해, 추출된 DNA를 중합효소연쇄반응(Polymerase Chain Reaction, PCR)에 사용하였다. 프라이머는 DPrbcL1 (5´-AAG GAG AAA THA ATG TCT-3´)과 DPrbcL7 (5´-AAR CAA CCT TGT GTA AGT CTC-3´)를 사용하였다(Jones et al., 2005). PCR 조건은 초기 변성(94℃, 3분)에 이어 94℃에서 50초, 52℃에서 50초, 72℃에서 1분 20초 조건을 36회 반복하였고, 최종 신장 조건은 72℃에서 15분 동안 수행하였다.
PCR 산물은 1% agarose gel (Bioneer, Korea)에서 100 V로 40분간 전기영동한 후, UV-transilluminator로 이동하여 DNA band 형성을 확인하였다. 증폭이 확인된 PCR 산물은 Cosmogentech (Seoul, Korea)에 의뢰하여 Sanger 염기서열 분석법으로 염기서열을 결정하였다. 확보된 염기서열은 GenBank 데이터베이스를 기반으로 BLAST 검색을 통해 유사 종을 확인하였으며, 계통수 작성을 위해 선정된 분류군의 염기서열과 함께 Clustal W (Thompson et al., 1994)를 사용하여 정렬하였다.
계통발생학적 분석 전 최적의 염기서열 치환 모델은 GTR+G+I로 설정하였다(Stepanek and Kociolek, 2016). 계통수는 Maximum Likelihood (ML)와 Bayesian inference (BI) 방법을 이용하여 각각 RAxML v.8.2 (Stamatakis, 2014) 및 BEAST v1.10.4 (Bouckaert et al., 2014) 소프트웨어로 분석하였다. ML 분석은 GTR+G+I 모델 하에서 1,000회 bootstrap 분석을 수행하였고(Felsenstein, 1985), BI 분석은 사후분포 조사를 1,500,000회 반복하여 수행하였다.
3. 결 과
3.1 형태학적 특징
Tetramphora sulcata (Brébisson) Stepanek & Kociolek, 2016 [Figs. 2A-I]
참고 문헌: Stepanek and Kociolek(2016), p. 6-7, figs. 22-34.
기재문: 엽록체는 Mereschkowsky의 분류에 따른 반원형 돌말류 엽록체 유형 중 제8형에 해당하며, 등 부분과 배 부분에 각각 두 개씩, 총 네 개의 엽록체가 큰 피레노이드(pyrenoid)와 연결된 형태로 관찰되었다(Fig. 2A). 세포는 중앙부가 넓고 정축 양 끝으로 갈수록 좁아지는 반타원형이다(Figs. 2A, C). 점무늬열(striae)은 정축 방향으로 길게 배열된 선형의 망목이 한 줄로 배열되어 있다(Fig. 2D). 등 부 가장자리는 아치형이며, 배부는 세포의 양 끝이 등 쪽을 향해 굽은 형태를 보인다. 등 줄은 정축 끝까지 연장되며 완만한 호 형태를 나타낸다(Figs. 2B, D, E). 배부 중앙에는 점무늬열이 없는 fascia 구조가 존재한다(Fig. 2E, 화살표). SEM 관찰 결과, 세포의 중앙부 내외부에서 두꺼운 규산질 구조가 확인되었고(Figs. 2F, H, 화살표), 정축 끝에서 등 줄이 안쪽으로 말려 있으며, 내부에는 Helictoglossa 구조가 존재하였다(Fig. 2I). 세포 크기는 길이 35–39 μm, 폭 8–9 μm이며, 10 μm당 점무늬열 밀도는 등부에서 19개, 배부에서 20개가 관찰되었다. 8-9 μm이다. 10 μm 당 점무늬열 밀도는 등부에서 19개, 배부에서 20개가 관찰된다.
국내 분포: 2021년 5월 경상북도 포항시 북구 청하면 청진2리(36°10'30"N 129°23'43"E), 자갈
세계 분포: Biscayne Bay, Bayfront Park, Miami-Dade County, Florida, USA.
Óbidos Lagoon, Óbidos, Portugal.
Neum Bay, Neum, Bosnia and Herzegovina.
Dalyan Iztuzu Beach, Dalyan, Muğla, Turkey.
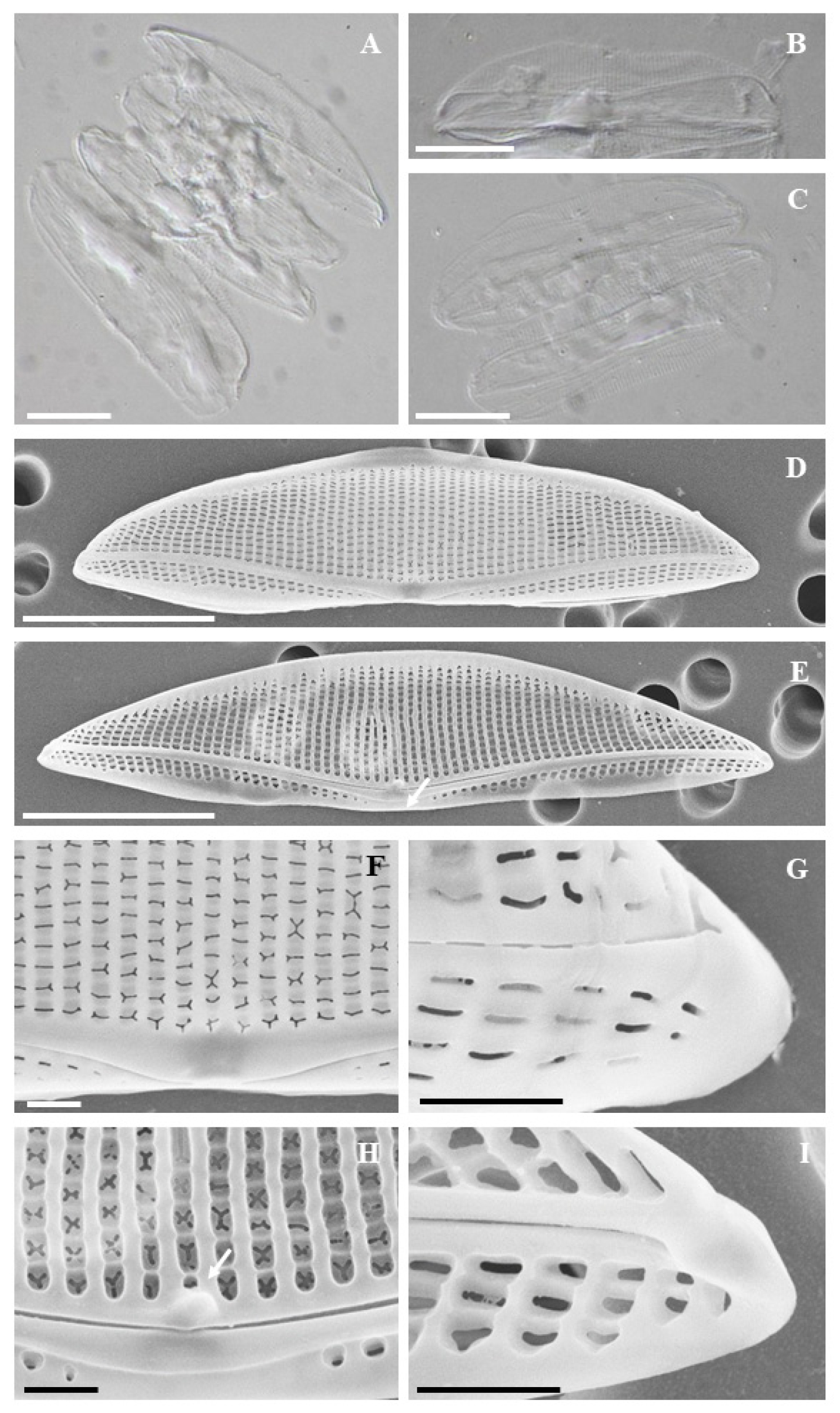
Fig. 2.
(A-C) Light microscopy, (D-I) SEM micrographs, (D) External valve view, (E) Internal valve view, arrow fascia (F) External valve center, (G) Details of external apex and raphe end, (H) Internal valve center, arrow projection, (I) Details of internal apex and raphe end, Scale bar = 10 μm (A-E), 1 μm (F-I).
3.2 계통발생학적 분석
Tetramphora 속의 염기서열 비교 분석에는 rbcL 유전자의 1개 marker alignment가 사용되었으며, 시퀀스 길이는 1,100 bp 이상으로 설정되었다. GenBank의 Tetramphora 종들과 비교한 결과, 연구 대상 종은 T. sulcata와 염기서열 유사도 100%로 일치하였다. 계통수는 Maximum Likelihood 및 Bayesian 분석을 통해 작성되었으며, 총 4종의 Tetramphora 종과의 계통학적 관계를 조사하였다(Fig. 3). 그 결과, 연구 대상 종은 T. sulcata와 동일한 계통군에 속하는 것으로 나타났다.
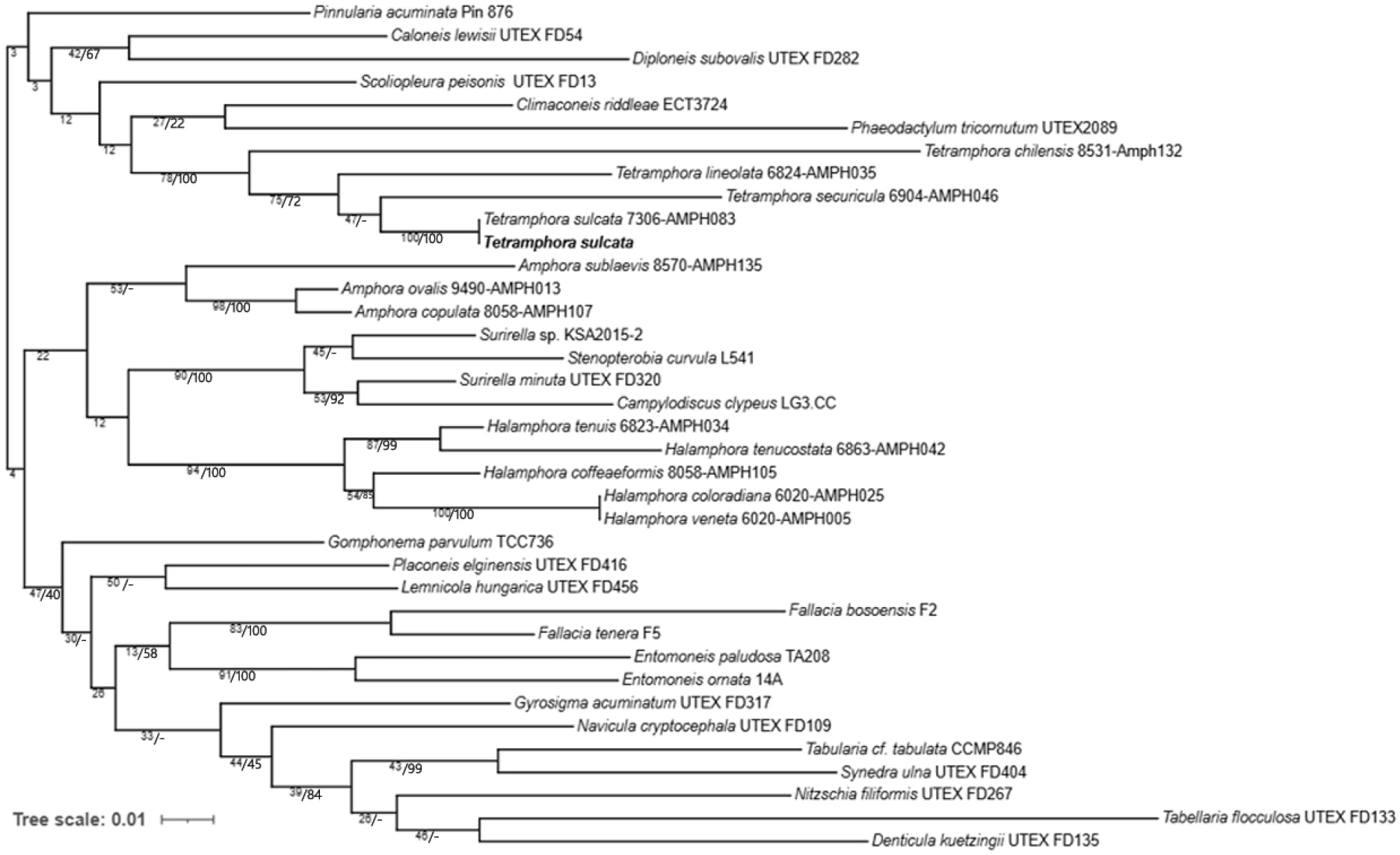
Fig. 3.
Molecular phylogenetic tree obtained from ML and BI analysis based on rbcL gene showing the phylogenetic positions of Tetramphora. Node support is positioned above or below the branch and is given as ML boostrap value/Bayesian posterior probability (as a percent). Node support (-) indicates discordance between the ML and Bayesian estimations.
4. 고 찰
전 세계적으로 Tetramphora 속은 12종이 보고되어 있으며, 국내에서는 Tetramphora rhombica,T. decussata, T. ostrearia, T. intermedia 등 4종이 기록되어 있다. 본 연구에서는 국내 저서 환경에 서식하는 Tetramphora 속의 형태 및 유전자 정보를 분석하였다. 광학현미경과 주사전자현미경을 이용한 형태 분석 결과, 본 연구 대상 종은 국내에서 보고된 T. intermedia와 valve 형태 및 raphe의 휘어진 정도에서 명확한 차이를 나타내었다. 특히, 크기 측면에서 본 종은 길이 35–39 μm, 폭 8–9 μm로, T. intermedia의 길이 68–114 μm, 폭 14–18 μm에 비해 더 작았다. 기존 종들과 비교해 보면, T. decussata는 등 쪽 중앙부에 두꺼운 규산질이 존재하며, 길이 75 μm, 폭 13–14 μm이다(심, 1994). T. rhombica는 길이 130–260 μm, 폭 21–50 μm로 훨씬 크고 넓은 형태를 보인다. T. ostrearia는 등 쪽 중앙부에 두꺼운 규산질이 있으며, 길이 40 μm, 폭 7 μm이다(Park and Koh, 2012).
유전자 분석 결과, 연구 대상 종은 기존에 보고된 Tetramphora sulcata와 매우 유사한 계통발생학적 위치를 나타내었다. 국내에서 출현한 T. sulcata는 폭이 다소 넓다는 점(Table 1)에서 기존 보고 종과 차이를 보였지만, 길이, stria 밀도 및 세부 형태 등에서 일치하였다(Stepanek and Kociolek, 2016).
Table 1.
Morphologic comparison of Tetramphora with related species. 1: Valve shape, 2: Raphe shape, 3: Ventral fascia, (+): present, (-): absent, n.d: no data
| species |
Length (㎛) |
Width (㎛) |
Dorsal Stria in 10 ㎛ |
Ventral Stria in 10 ㎛ | 1 | 2 | 3 | ref. |
| T. sulcata (KNUY-21121) | 35-39 | 8-9 | 19 | 20 | semi-lanceolate | Dorsally | + | This study |
| T. sulcata | 35-58 | 5.5-7.5 | 18-19 | - | semi-lanceolate | Dorsally | + | Stepanek and Kociolek, 2016 |
| T. securicula | 55-75 | 8-10 | 16 | - | semi-lanceolate | Dorsally | - | Stepanek and Kociolek, 2016 |
| T. chilensis | 20-42 | 4.5-6 | 20-23 | 26-30 | semi-lanceolate | Dorsally | + | Stepanek and Kociolek, 2016 |
| T. lineolata | 25-48 | 5.5-8 | 21-23 | - | semi-lanceolate | Dorsally | - | Stepanek and Kociolek, 2016 |
| T. intermedia | 68-114 | 14-18 | 14-18 | n.d | semi-lanceolate | Dorsally | - | Park et al., 2017 |
| T. rhombica | 130-260 | 21-50 | 10 | n.d | semi-lanceolate | Dorsally | n.d | Stepanek and Kociolek, 2016 |
| T. decussata | 75-85 | 13-14 | 16-18 | n.d | semi-lanceolate | Dorsally | n.d | 심, 1994 |
| T. ostrearia | 40 | 7 | 12 | n.d | semi-lanceolate | Dorsally | - | Park and Koh, 2012 |
따라서 국내에는 기존에 보고된 4종 외에 T. sulcata를 포함하여 총 5종의 Tetramphora 속 종이 분포하는 것으로 판단된다.
