

1. 서 론
2. 재료 및 방법
3. 결 과
3.1 야광충 개체수
3.2 먹이생물 조성과 개체수
4. 토 의
4.1 야광충 출현 개체수 및 분포특성
4.2 야광충 개체수와 식물플랑크톤
4.3 야광충과 먹이생물 다양성
1. 서 론
야광충(Noctiluca scintillans) (Ehrenberg) Kofoid & Swezy은 세포직경이 200~800 ㎛에 이르는 무각(unarmoured)의 해양 부유성 와편모류이다(Fonda-Umani et al., 2004; Fukuda and Endoh, 2006). 엽록체가 없고 세포질(cytoplasm)이 색깔이 없는 무색이며, 섭식능력이 있다. 수 많은 식포(food vacuoles)가 세포질 내에 존재하여 강한 부유능력이 있어서, 해류에 떠서 쉽게 이동하지만 활발히 움직일 수는 없다. 그리고 야간에 선박이나 파도에 부딪혀 빛을 발산하는 생물발광(Bioluminescent) 생물이기도 하다(Fukuda and Endoh, 2006). 야광충은 적조원인종으로 널리 알려진 생물이며, 그 분포범위도 온대해역, 아열대해역과 열대해역에 이르는 매우 넓은 해역이다(Uhlig and Sahling, 1990; Fonda-Umani et al., 2004; Miyaguchi et al., 2006; Mohamed and Mesaad, 2007; Mohanty et al., 2007; Mcleod et al., 2012; Tsai et al., 2018). 야광충 대발생을 촉발하는 요소는 흔히 물리적인 기작(바람, 해류, 용승지역, 전선역 등)에 의해 야광충이 모이면(Uhlig and Sahling, 1990; Huang and Qi, 1997; Dela-Cruz et al., 2002; Fonda-Umani et al., 2004), 적절한 수온, 잔잔한 환경과 먹이농도가 대발생을 야기한다(Huang and Qi, 1997; Dela-Cruz et al., 2002; Fonda-Umani et al., 2004; Miyaguchi et al., 2006; Mohamed and Mesaad, 2007). 야광충의 개체수 증가요인이 매우 다양하지만, 많은 경우에 높은 농도의 먹이생물이 존재할 때, 특히 규조류가 우점하는 조건하에서 높은 개체수의 야광충이 관측되었다(Kiørboe et al., 1998; Dela-Cruz et al., 2002, 2003; Mohamed and Mesaad, 2007; Mcleod et al., 2012; Tsai et al., 2018).
야광충은 식세포섭식(phagotrophic) 생물이며, 주요 먹이생물이 식물플랑크톤인 규조류(diatoms)로 알려져 있으나, 다양한 먹이스펙트럼(박테리아, 분변립, 잔재물질, 원생생물, 요각류, 요각류 알, 어류)을 갖는 것으로 알려져 있다(Daan, 1987; Nakamura, 1998a, 1998b; Quevedo et al., 1999; Kiørboe, 2003; Yilmaz et al., 2005; Escalera et al., 2007; Suzuki et al., 2013; Tsai et al., 2018). 야광충 대발생은 빨간색의 토마토 수프와 같은 성상을 나타내어 용존산소 결핍과 양식어류와 해양무척추생물의 폐사로 이어진 사례가 있다(Huang and Qi, 1997; Fonda-Umani et al., 2004). 야광충이 독성물질을 생산하지 않더라도, 독성수준의 암모니아를 축적하고 주변해수에 배출하여 치사요인(killing agent)으로 작용하는 것으로 알려져 있다.
야광충의 분포특성 연구는 대부분 연안역과 항만환경에서 수행되었다(Fonda-Umani et al., 2004; Miyaguchi et al., 2006; Yoo et al., 2006; Mohamed and Mesaad, 2007; Kang, 2010; Baek et al., 2011). 국외 연안역에서 관측된 최대개체수가 단위체적당 100,000개체(Fonda-Umani et al., 2004; Miyaguchi et al., 2006; Mohamed and Mesaad, 2007)였던 반면, 국내연안에서는 단위체적당 10,000개체 범위였다. 이처럼 야광충은 연안역에서의 연구에 비해 외해역에서 매우 드물며, 동물플랑크톤 군집특성 연구 시 분석된 정도에 그쳤다. 동해 독도 주변해역에서 관측된 야광충 개체수는 1997년과 1998년 하계에 3~93 inds.m-3이었고(Association for Research and Conservation of Dokdo, 1999), 2002년에 춘계와 추계에 평균 1~20 inds.m-3이었다(Kang et al., 2002). 2006~2015년 하계에는 독도 주변해역에서 3,990~13,818 inds.m-3의 범위에서 점차 증가하였다(Kang et al., 2016).
현재까지의 야광충의 개체군 변동과 분포특성 연구에서, 환경요인(수온, 염분)과 생물학적 요인(주로 엽록소-a 농도)간의 상호관계가 주요 연구주제인 반면, 대발생이 관찰되었을 때 야광충과 먹이생물 간의 포식-피포식 관점에서 야광충 식포(food vacuole)에 어떤 종류의 생물이 존재하는지의 직접적인 섭식 정보는 알려진 바가 없다. 식포 내의 먹이생물의 정성, 정량적 정보는 야광충 먹이생물의 직접적 정보가 될 뿐만 아니라, 포식자로서 먹이생물 군집의 하향조절(top-down control)특성과 야광충 개체군 크기에 대한 생태학적 이해도를 높일 수 있다. 본 연구는 독도 주변해역에서 2014년 춘계에 관측된 야광충의 대발생 시 식포 내에 존재하는 생물의 종류와 양을 파악하여, 야광충의 먹이 섭식특성을 이해하고자 하였다.
2. 재료 및 방법
야광충 식포 내 먹이생물 관찰을 위한 채집은 2014년 5월 1일에 독도 주변 6개 정점에서 수행되었다(Fig. 1). 독도 주변해역 정점에서의 야광충 시료는 한국해양과학기술원의 수행과제(독도의 지속가능한 이용연구)의 일환으로 독도 주변해역에 분포하는 동물플랑크톤의 군집 및 생물량 조사가 수행된 춘계 조사기간(2014년 4월 29일~5월 3일) 중 획득되었다. 조사 정점은 울릉도 동부연안에서 독도를 가로질러 동부 정점에 이르는 총 6곳(16, 20, 30, 47, 45, 50)이었다(Table 1, Fig. 1). 한국 해양과학기술원 해양조사선 “이어도 호”를 이용하여 매 정점에서 채집에 앞서 CTD (SBE 911 plus)를 내려 수온과 염분의 수직분포를 확인한 후, 표층혼합층에서 수직채집하였고, 표층혼합층 깊이는 수온변화범위가 표층 수온의 1℃ 이내인 곳까지 감안하여 결정하였다(Table 2). 동물플랑크톤 채집은 표준네트(직경 60 ㎝, 망구크기 200 ㎛)를 이용하였고, 정선 상태에서 목표수심까지 네트를 내린 후, 50 ㎝/sec의 속도를 유지하여 표층까지 예인하였다. 여과해수 부피의 정량을 위해 유량계(Hydro-Bios Kiel 438110: back-run stop)를 표준네트 입구에 부착하여 회전수를 기입하였다. 채집시료는 선상에서 채집통을 주의하여 세척한 후 시료병에 옮겨담은 후, 중성 포르말린으로 최종농도 5%가 되도록 고정하였다. 실험실로 운반 후 시료병 내의 내용물이 잘 섞이도록 충분히 흔든 후 10-30 mL을 분취하여 계수판(Bogorov counting tray)에 옮겨 해부현미경(SteReo SV8, Zeiss)하에서 ×50~80배의 배율에서 야광충을 동정 및 계수하였고, 식포 내 먹이생물의 유․무를 기준으로 구분하여 계수하였다. 연구기간 중 야광충 식포 내에 존재하는 먹이생물의 정량분석을 위해 시료병에서 분취한 일정부피(10-30 mL)내에서 관찰하여 먹이를 섭식한 야광충을 골라낸 후, 관찰한 부피 내의 먹이생물 개수와 종조성을 확인하여 단위체적 당 총 개수(inds·m-3)로 환산하였다. 대부분 야광충 100개체 정도를 관찰했고, 먹이생물 출현율이 가장 낮았던 정점 50 (24%)에서는 최대 900개체까지 관찰하였다. 야광충에 섭식된 생물은 광학현미경(Axioplan 11, Zeiss)하에서 ×400배의 배율로 동정하였고, 네트 플랑크톤 이상(>20 ㎛)의 생물을 대상으로 하였다. 야광충 사진은 영상분석 프로그램(AxioCam ICc 3, ZEN 2011 programme, SteREO Discovery, V8; Zeiss)으로 촬영하였고, 야광충 식포 내의 먹이생물사진은 ×400배의 광학현미경하에서 영상분석 프로그램(AxioCam ICc 3, ZEN 2010 program; Axio Imager A2; Zeiss)으로 촬영했다. 식포 내의 다양한 생물(식물플랑크톤, 원생생물 및 동물플랑크톤)은 Shim(1994)과 Chihara and Murano(1997) 그리고 Yoo(1995)를 참조하여 분석하였다.
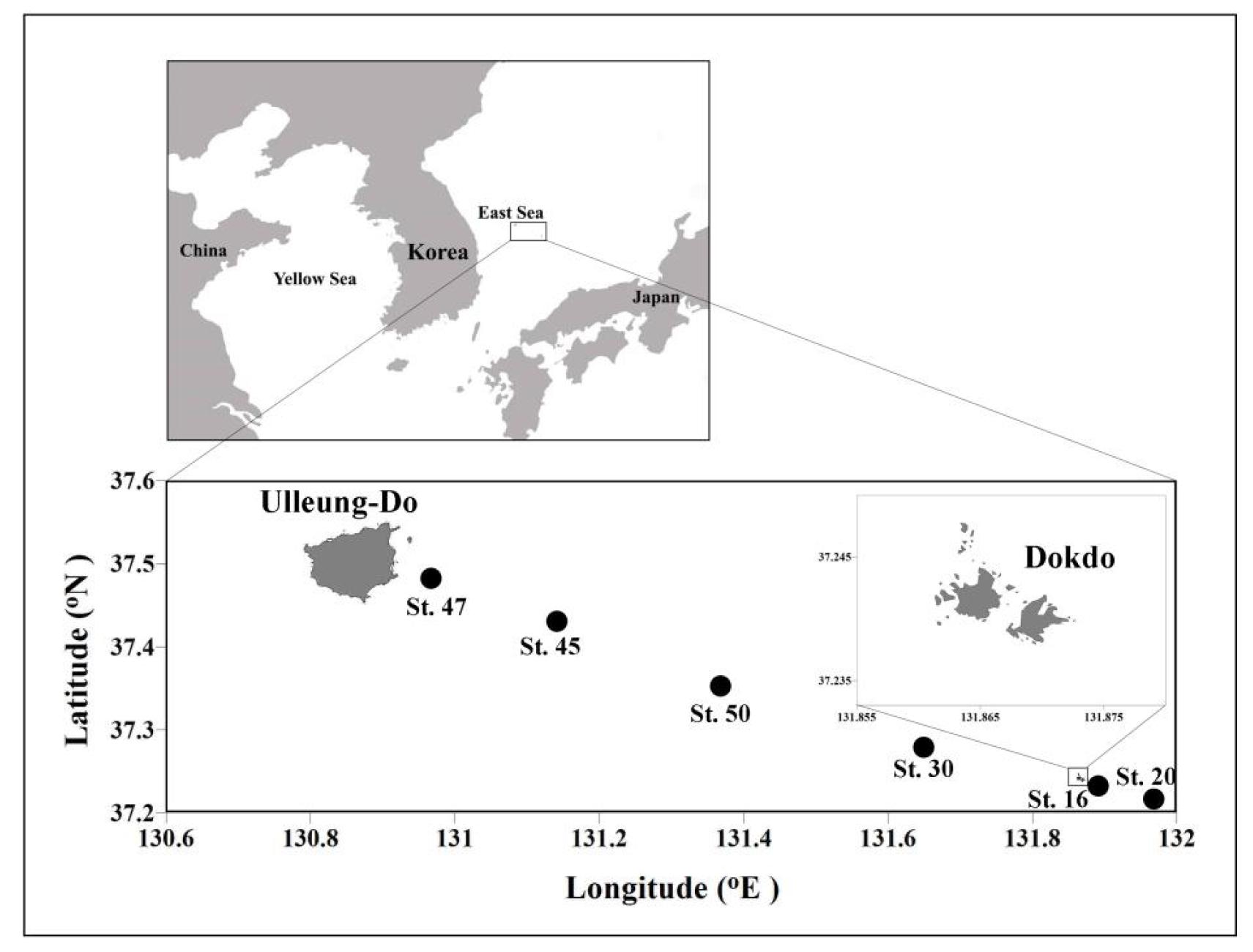
Table 1.
Location for sampling of Noctiluca scintillans and the preyed items in the food vacuole of the species around Dokdo in the East Sea in spring 2014
| Station | Latitude | Longitude |
| 16 | 37"13·35.0“ | 131"53·49.0“ |
| 20 | 37"08·02.8“ | 132"03·36.0“ |
| 30 | 37"17·02.4“ | 131"38·24.0“ |
| 45 | 37"23·29.0“ | 131"14·24.0“ |
| 47 | 37"27·02.9“ | 131"00·46.8“ |
| 50 | 37"19·51.8“ | 131"25·19.1“ |
3. 결 과
3.1 야광충 개체수
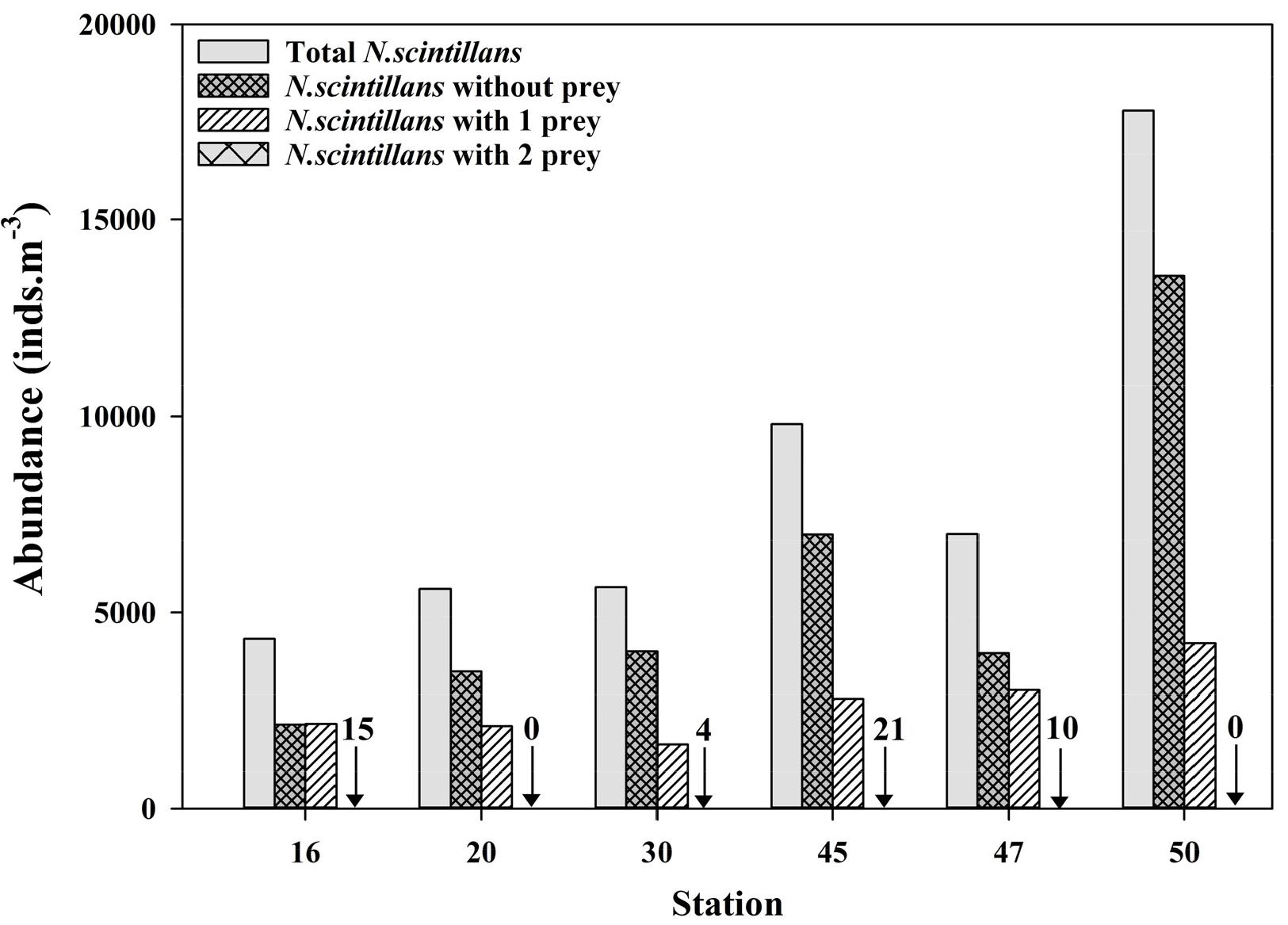
야광충은 연구선(이어도 호)에 승선하여 울릉도 동쪽에서 독도를 잇는 장기모니터링 정점(6개)의 표층혼합층에서 채집하여 분석하였다. 출현한 야광충은 4,328~17,791 inds.m-3의 범위에서 정점 50에서 가장 높았고, 정점 30에서 가장 낮았다. 야광충 식포 내에 먹이생물이 존재하는 개체수는 1,642~3,030 inds.m-3(출현율: 24~50%)이며, 먹이생물이 없는 야광충 개체수는 2,149~13,577 inds.m-3(출현율: 50~76%)이었다(Fig. 2). 독도 주변해역에서 식포 내에 먹이생물이 종조성과 상관없이 1 개체가 있는 야광충은 1,638~3,030 inds.m-3 범위에서 출현했고, 2 개체가 있는 야광충은 0~21 inds.m-3범위에서 출현하였으며, 그 이상의 개체를 포함한 야광충은 없었다(Fig. 2).
3.2 먹이생물 조성과 개체수
채집된 야광충 식포 내 먹이생물은 분명한 형태를 갖는 것을 대상으로 분석하였고, 원래의 형태를 알아보기 힘든 정도로 소화된 것은 제외하였다. 야광충 식포 내에서 관찰된 먹이생물은 식물플랑크톤(규조류와 와편모조류), 원생생물, 요각류, 어란, 치어, 분변립(fecal pellet) 그리고 소나무의 꽃가루(pollen)가 확인되었다. 규조류는 중심형 규조목이 대부분이었고, 와편모조류는 적조원인종으로 알려진 Heterocapsa sp., Prorocentrum spp. 그리고 Protoperidinium sp.이었다. 원생생물은 유종섬모충류가 관찰되었고, 동물플랑크톤은 요각류, 어류플랑크톤은 어란과 치어가 관찰되었다. 그리고 동물플랑크톤의 것으로 판단되는 분변립이 관찰되었고, 소나무의 꽃가루까지 관찰되었다(Table 3, Fig. 3).
Table 3.
Abundance of preyed organisms in the food vacuole of Noctiluca scintillans collected in the surface mixed layer around Dokdo in the East Sea in spring 2014
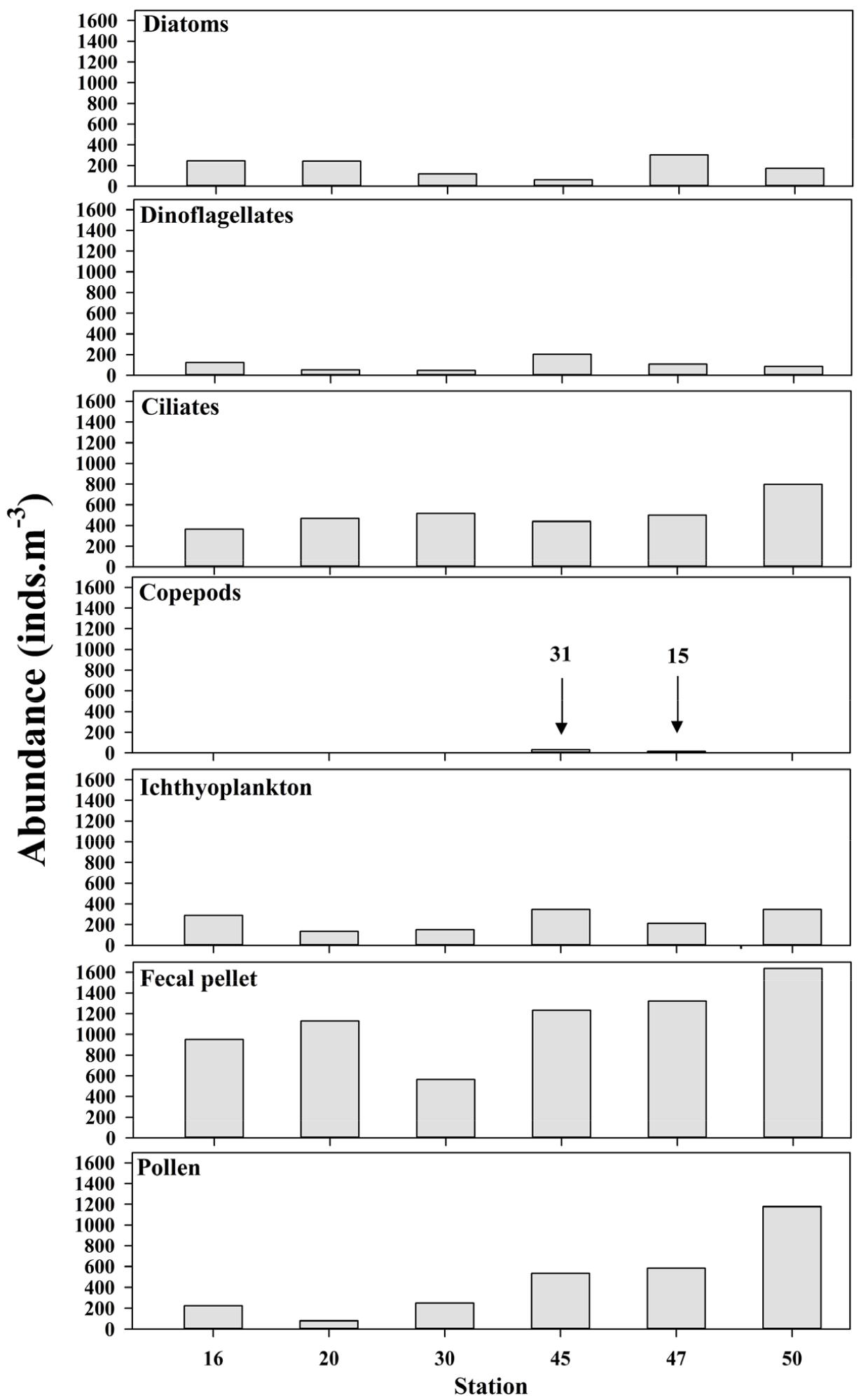
식물플랑크톤 중에서 규조류인 Thalassiosira sp.가 와편모조류 Protoperidinium sp.보다 야광충 식포 내에서 더 많이 관찰되었다. 그리고 원생생물, 어란 그리고 분변립이 야광충 식포 내에서 매우 높은 개체수를 나타냈으며, 이는 다른 식물플랑크톤과 동물플랑크톤에 비해 매우 높은 값이었다(Table 3).
독도 6개 정점에서 먹이생물 분류군 별 평균 출현율의 경우, 분변립이 가장 높았고(43%), 원생생물이 19%, 꽃가루가 18%를 차지했고, 어란과 치자어가 9%, 규조류가 7%, 그리고 와편모조류가 4%를 나타냈다(Table 3, Fig. 3).
식포 내에 먹이생물을 포함한 야광충 사진들을 모았으며, 그 종류는 Table 3에 기록된 생물들로 구성되었다. 특히 지각류의 경우, 먹이생물 정량분석 시에 출현하지 않았으나, 별도로 추가 분석했을 때 관찰되었으며, Evadne nordmanni로 분석되었다(Fig. 4).
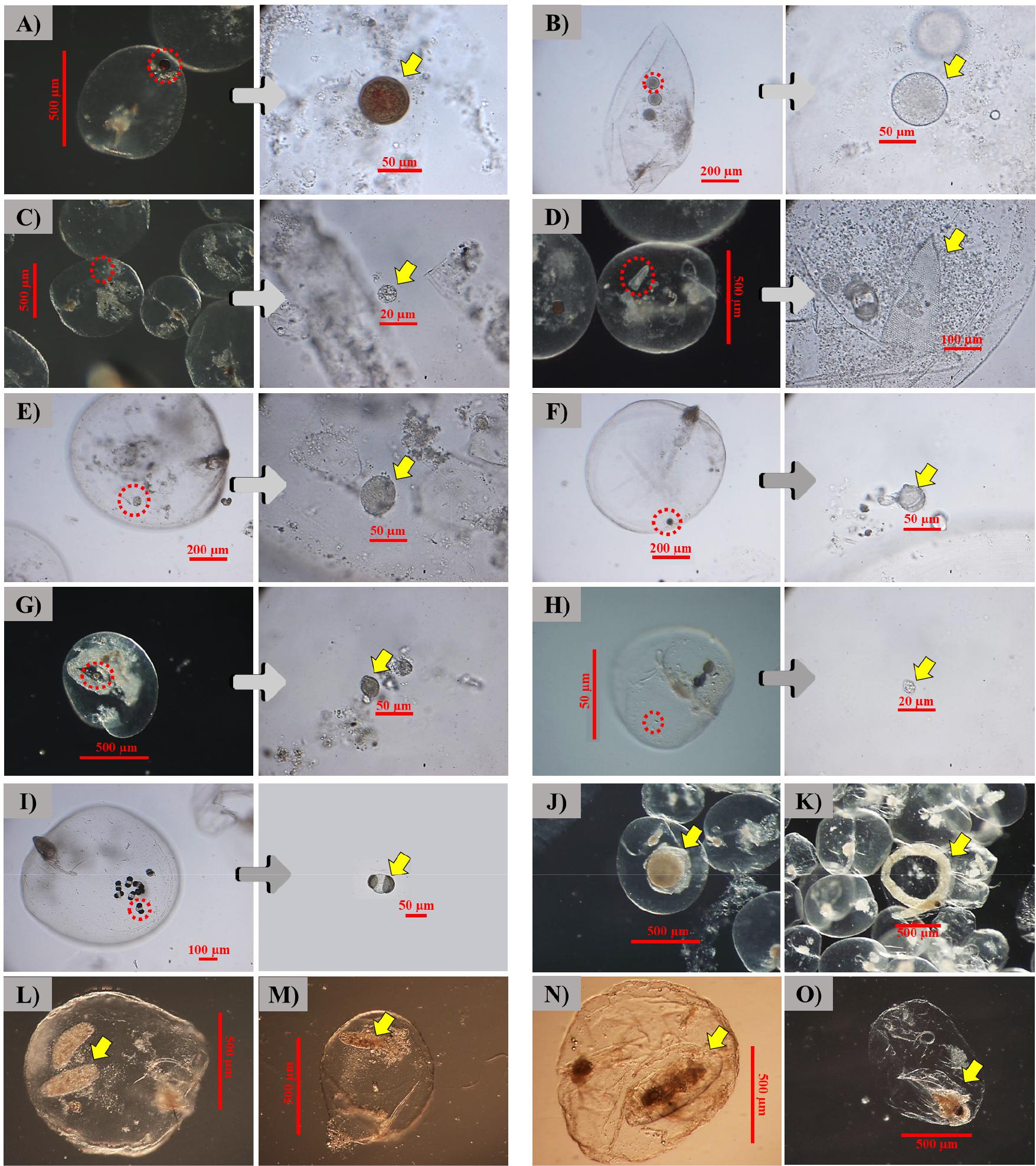
Fig. 4.
Food items found in the vacuole of Noctiluca scintillans sampled around Dokdo in the East Sea in spring 2014. A) Thalassiosira eccentrica, B) Coscinodiscus radiatus, C) Thalassiosira pacifica, D) Parafavella sp., E) Prorocentrum micans, F) Protoperidinium sp., G) Prorocentrum sp., H) Heterocapsa sp., I) pollen, J) fish egg, K) fish larvae, L-M), fecal pellets, N) copepods, and O) Evadne nordmanni.
4. 토 의
4.1 야광충 출현 개체수 및 분포특성
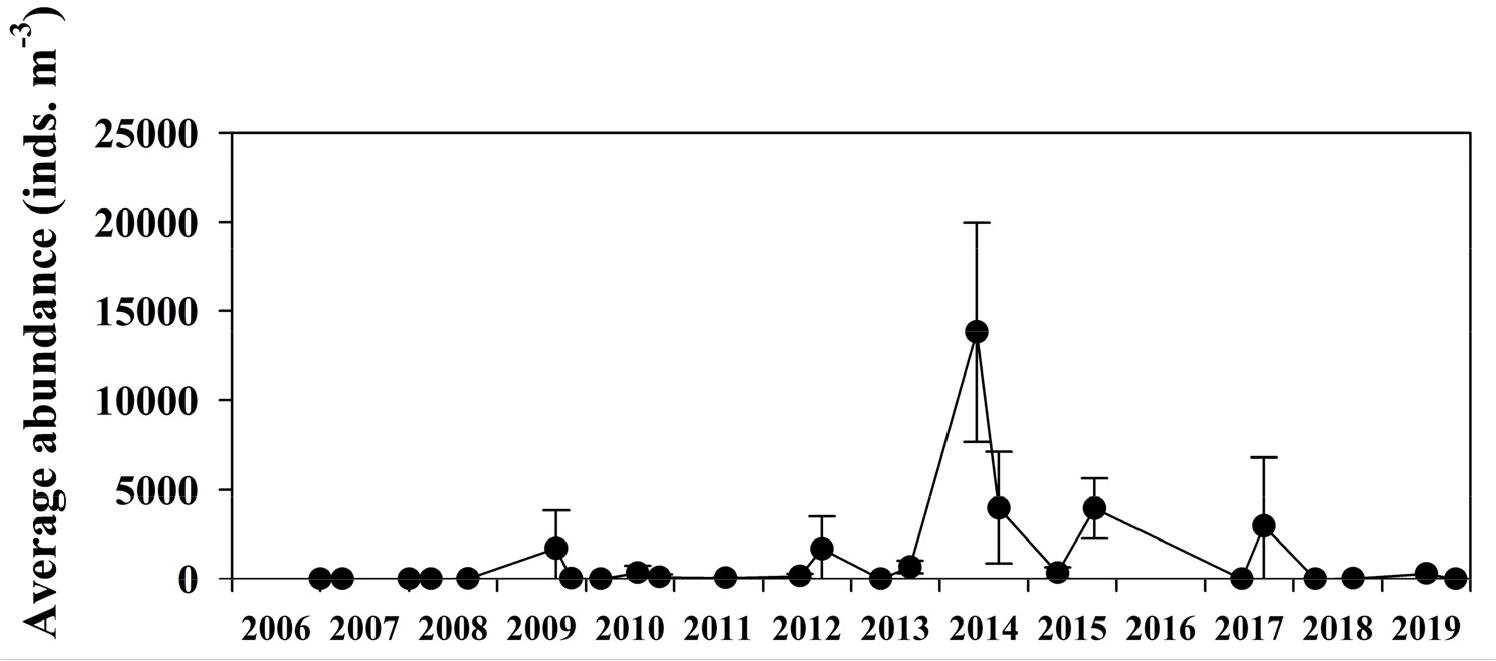
본 연구기간에 울릉도 동부에서 독도 동부를 잇는 6개 정점(4,328~17,791 inds.m-3)에서 출현한 개체수는 우리나라 연안역에서 관측된 최대개체수 [인천연안: 7.7×104 inds.m-3, 주요항만: 41,753 inds.m-3]와 유사한 범위이거나 비교할 수 없이 높았다(Yoo et al., 2006; Kang, 2010). 일반적으로 야광충은 연안에서 주로 관측되는 것으로 알려져 있으나(Harrison et al., 2011), 외양에서 출현한 개체수가 연안역에 비해 높았거나 유사한 사례는 현재까지 없었다. 동해 외해역에서 야광충이 출현한 기록은 독도 주변해역, 동해 전선역, 울릉분지 그리고 동해 서남 해역 등이 주를 이루었다. 독도 주변해역에서 야광충의 출현기록은 1997년 7월에 0~59 inds.m-3, 1998년 7월에 26~93 inds.m-3의 범위였다(Association for Research and Conservation of Dokdo, 1999). 1999년 9월에는 5~30 inds.m-3, 2000년 5월에는 0~3 inds.m-3의 범위에서 출현하였다(Kang et al., 2002). 2006년에 시작된 “독도의 지속가능한 이용연구” 과제에서 2011년까지는 주로 여름철에 출현했고(최대 개체수 3,990 inds.m-3), 2014년 봄철의 최대개체수(0~100 m)는 평균 13,818 inds.m-3를 나타냈다(Kang et al., 2016). 독도종합시스템에서 나타난 장기간 야광충 개체수 변동자료(2006년~2019년)를 보면, 2014년 봄철의 야광충 개체수가 가장 높았으며, 이는 본 연구결과에 나타난 야광충 개체수가 장기간동안 매우 높았던 값임을 의미하였다(Fig. 5, www.dokdo.re.kr).
동해 서남부해역에서는 최대 311 inds.m-3가 출현했고(Shim and Lee, 1986), 동해 전선역에서는 최대 224 inds.m-3를 나타냈다(Park and Choi, 1997). 난수성 소용돌이 구조가 관측되었던 울릉분지에서 가장 높은 개체수(200 inds.m-3)가 소용돌이 구조 중심부에서 관측된 바가 있다(Kang et al., 2004). 따라서 현재까지 보고된 동해에서의 야광충 개체수에 비해, 본 연구시기에 관측된 야광충의 개체수가 예외적이었음을 알 수 있다. 비록 이 개체수는 아드리아 해와 사가미 만에서 관측된 최대개체수인 142,477 inds.m-3와 610,000 inds.m-3보다 낮으나(Fonda-Umani et al., 2004; Miyaguchi et al., 2006), 동해 외해역에서의 출현개체수라는 관점에서 매우 주목할 만하다.
4.2 야광충 개체수와 식물플랑크톤
야광충 개체수의 증가에 영향을 끼치는 조건은 환경(bottom-up regulation)과 생물학적 조건(top-down control) 관점에서 이해할 수 있다. 환경조건은 야광충 성장에 미치는 결정적 환경요인(수온, 염분 등)의 특성과 해양학적 과정(용승, 소용돌이 구조, 해류, 바람 등)으로 설명될 수 있으며, 생물학적 조건은 야광충이 해양환경에서 섭식가능한 생물의 조성과 양으로 설명될 수 있다(Uhlig and Sahling, 1990; Huang and Qi, 1997; Dela-Cruz et al., 2002; Fonda-Umani et al., 2004). 대다수의 연안역과 외해역 현장연구들은 야광충의 개체수와 분포특성을 결정하는데 축적기작의 중요성(바람과 해류)을 언급하고 있으며(Kang et al., 2004; Miyaguchi et al., 2006), 야광충의 개체수 증가가 각 지역별로 특징적인 해양학적 과정의 결과로 나타난 먹이생물의 증가와 관련있음을 보고하였다(Dela-Cruz et al., 2002; Kang et al., 2004; Miyaguchi et al., 2006; Dela-Cruz et al., 2008; Mcleod et al., 2012; Sarma et al., 2019; Xiang et al., 2019). 특징적으로, 호주 동부연안에서 야광충의 개체수 증가 이전에 규조류가 우점하는 식물플랑크톤의 증가가 선행되며, 야광충의 식포 내에서(>80%) 주로 규조류(Thalassiosira partheneia)가 관찰되거나 된 바 있다(Dela-Cruz et al., 2002; Dela-Cruz et al., 2008). 그리고 브라질 연안외측의 용승 상승류(upwelling plume)에서 증가한 야광충 식포 내에서 분변립(fecal pellets)이 우점한 바가 있으며(Kiørboe, 2003), 아라비아 북부 해역에서는 비규조류 식물플랑크톤의 대발생과 입자성 유기탄소 물질이 야광충 대발생을 야기한 바 있다(Sarma et al., 2019; Xiang et al., 2019). 2006년 이후 2019년까지 독도 주변해역에서 가장 높은 개체수의 야광충이 관찰된 2014년 춘계에 평균 엽록소-a 농도가 가장 높았다(www.dokdo.re.kr). 비록 식물플랑크톤 군집의 정량 및 정성분석 자료가 없어서, 2014년 춘계에 관측된 높은 농도의 엽록소-a의 출현기작을 알 수 없으나, 상대적으로 높은 식물플랑크톤 생물량이 야광충의 높은 개체수와 관련이 있었던 기존사례와 유사하였다. 이처럼 먹이생물의 이용가능성은 야광충 개체수 변화에 미치는 중요한 영향요인 중 하나일 수 있다(Kiørboe et al., 1998; Dela-Cruz et al., 2002; Zhang et al., 2017; Kitatsuji et al., 2019).
4.3 야광충과 먹이생물 다양성
야광충은 먹이 선택성이 거의 없으며, 다양한 생물들을 섭식할 수 있다(박테리아, 분변립, 식물플랑크톤, 원생생물, 소형동물플랑크톤, 요각류, 요각류 알, 어류란, 치자어) (Huang and Qi, 1997; Kiørboe, 2003; Fonda-Umani et al., 2004; Zhang et al., 2017; Kitatsuji et al., 2019). 그래서 야광충은 생태계 내에서 섭식한 먹이생물의 군집 혹은 개체군을 조절할 수 있는 가능성이 있다. 앞서 언급된 바와 같이, 야광충 개체수가 증가하기 시작할 때 식물플랑크톤 중에 규조류가 주로 선택되었던 반면, 같은 식물플랑크톤인 와편모조류 현존량이 증가할 때, 야광충의 성장률이 감소하는 것으로 보아, 와편모조류는 규조류에 비해 덜 선호되는 먹이임이 중형폐쇄생태계(mesocosm) 실험에서 확인되었다(Zhang et al., 2017). 현장연구에서 규조류 대발생 후 야광충 증가가 빈번히 관찰되며(Kiørboe and Titelman, 1998; Nakamura, 1998b; Dela-Cruz et al., 2002; Zhang et al., 2017; Kitatsuji et al., 2019), 엽록소-a 농도와 야광충 개체수와의 높은 상관성이 보고된 바가 많았는데, 이는 식물플랑크톤에 대한 강한 섭식압을 의미한다(Huang and Qi, 1997). 그리고 규조류가 와편모조류에 비해 더 많이 섭식되는 이유로서, 규조류가 야광충 촉수 끝 부분의 끈적한 실 부분에 잘 잡히며, 운동성이 적은 것이 대표적인 것임이 실험을 통해 확인되었다 (Uhlig and Sahling, 1990; Kiørboe and Titelman, 1998).
해양환경에서 야광충의 높은 개체수가 관찰되었을 때, 식포 내에서 관찰된 먹이생물과 실험실 연구에서 수행된 야광충 식포 내에서 관찰된 먹이생물은 식물플랑크톤 외에 매우 다양하게 나타났다(Kimor, 1979; Daan, 1987; Shanks and Walters, 1996; Nakamura, 1998a; Dela-Cruz et al., 2002; Escalera et al., 2007; Mohamed and Mesaad, 2007; Mcleod et al., 2012; Drits et al., 2013; Suzuki et al., 2013; Tsai et al., 2018; Kitatsuji et al., 2019). Shanks and Walters(1996)는 워싱턴 San Juan Islands에서 채집된 해중설(marine snow)에 야광충이 모여있는 것을 관찰했고, 해중설의 끈적한 물질에 존재하는 와편모조류의 포자(cysts)가 야광충 식포에서 관찰되었다(60~80%). Nakamura(1998a)는 10 ㎛ 이상의 먹이생물 중 Chattonella antiqua와 Heterosigma akashiwo (Raphidophyceae)가 야광충의 성장에 의미있는 영향을 끼쳤음을 보고하였다. Nakamura(1998b)는 일본 세토 내만에서 야광충의 성장률이 규조류 대발생 시기에 최고였으며, 여름철 중형동물플랑크톤의 주요 구성원 중 하나임을 보고하였다. 호주 동남해안에서 규조류 대발생 시기에 관찰된 야광충의 80% 이상에서 먹이생물(주로 소형 중심형 규조류인 Thalassiosira partheneia)이 식포 내에서 관찰되었으며, 뒤 이어 관찰된 야광충 개체수의 증가에 영향을 끼쳤음을 알 수 있었다(Dela-Cruz et al., 2002). 스페인 해역에서 출현한 야광충 식포 내에서는 Dinophysis와 Pseudo-nitzschia속의 식물플랑크톤이 관찰되었고(Escalera et al., 2007), 홍해에서는 식포 내에서 규조류(Thalassiosira rotula and Pseudonitzschia sp.), 와편모조류(Dinophysis sp.와 Prorocentrum sp.) 그리고 녹조류(Dunaliella tertiolecta)가 관찰되었다(Mohamed and Mesaad, 2007). 일본 큐슈 서해외해역에서 관찰된 야광충의 식포 내에서는 규조류가 와편모조류에 비해 더 많았고(60~100%), 우상형 규조류보다 중심형 규조류(Chaetoceros and Bacteriastrum)가 더 많이 관찰되었다(Suzuki et al., 2013). 이는 긴 강모를 가진 사슬형 콜로니형태인 중심형 규조류가 우상형 규조류보다 야광충의 촉수에 더 쉽게 잡히기 때문일 것으로 제시하였다. 남극해에서 관측된 야광충의 40% 정도에서 Thalassiosira spp.가, 20% 정도에서 Guinardia striata가 관찰되었다(Mcleod et al., 2012). 흑해에서는 Neoceratium tripos와 N. furca 그리고 Alexandrium에 속하는 와편모조류의 일시성 포자(cyst)가 야광충 식포 내에서 관찰되었다(Drits et al., 2013). Matsu archipelago의 아열대 연안수역에서 출현한 야광충의 식포에서는 규조류(Thalassiosira spp.)가 관찰되었다(Tsai et al., 2018). 일본 세토 내만에서 야광충의 빠른 성장이 관찰되었을 때, 야광충 식포 내에서 규조류인 Chaetoceros spp., Thalassionema spp., Thalassiothrix spp., Coscinodiscus spp., Thalassiosira spp.와 Stephanopyxis spp.이 관찰되었고, 규조류 생물량의 33.6%의 감소를 야기하였다(Kitatsuji et al., 2019). 한편, 흑해에서 관찰된 야광충 식포 내에서 요각류 Calanus euxinus의 알 생산력의 2-9%가 관찰된 바가 있다(Drits et al., 2013).
본 연구기간 독도 주변해역에서 출현한 총 야광충 식포 내에서 먹이생물이 평균 32% (24~50%)가 출현했고, 관찰된 먹이생물 조성은 식물플랑크톤의 경우 중심형 규조류와 와편모조류가 각각 7%와 4%의 출현율을 나타냈으며, 종 조성 측면에서 기존 연구결과와 유사하였다(Table 3). 그러나 분변립의 압도적인 출현율(43%)과 꽃가루의 출현(18%)이 매우 특징적이었다. 기존에 알려진 섭식먹이생물의 조성 측면에서 식포 내에서 꽃가루의 출현은 매우 이례적이며, 출현율 측면에서 분변립의 높은 갯수는 매우 특이적이다. 그러나 운동성이 거의 없는 먹이의 경우 섭식율이 높다는 측면에서 분변립과 꽃가루의 높은 출현율은 이해할 만하다(Kiørboe and Titelman, 1998). 그러나 규조류에 비해 훨씬 더 많이 섭식된 점은 기존 연구결과와 매우 다르다.
해양환경에서 식물플랑크톤 현존량이 낮은 경우 원생생물, 작은 크기의 요각류와 요각류 알을 섭식하여 기아(starvation)를 피하고, 같은 먹이를 취하는 소형 및 중형 동물플랑크톤과의 경쟁을 회피하며, 동시에 동물플랑크톤 포식자와 경쟁을 하는 경우도 있다(Zhang et al., 2017). 식물플랑크톤이 거의 없을 때에는, 와편모조류보다 요각류 알이 더 선호되었으며, 최고 34%까지 차지하였다. 남부 캘리포니아 내만에서 관찰된 야광충 식포 내에서 요각류 Acartia tonsa의 알이 관찰된 바가 있으며, 이와 비슷한 크기의 알을 생산하는 다른 요각류 개체군에 영향을 끼칠 수 있다(Kimor, 1979). 한편, 야광충이 지중해와 흑해를 잇는 Marmara해역에서 초미소플랑크톤과 미소플랑크톤을 섭식하여 경쟁관계에 있는 요각류 A. clausi의 먹이이용가능성을 감소시킨 바가 있다(Yilmaz et al., 2005). 그리고 북부 아드리아 해에서 야광충이 소형동물플랑크톤(microzooplankton)을 섭식하여, 식물플랑크톤에 대한 소형동물플랑크톤의 섭식압이 16%까지 감소한 예가 있다(Fonda-Umani et al., 2004). Mcleod et al.(2012)는 야광충 대발생이 관찰된 곳에서 낮은 개체수의 요각류가, 야광충이 감소한 곳에서 요각류 개체수가 증가한 관측에 근거하여, 야광충이 요각류 군집과 잠재적으로 경쟁한다고 보고하였다. 한편, 스페인의 중앙 Cantabrian 연안역에서 급증가한 야광충이 요각류(A. clausi) 알을 총 량의 70%까지 직접 섭식하여 유생의 가입(recruitment) 잠재성을 상당히 낮춰 요각류 개체군 변동에 영향을 줄 수 있음을 보고하였다(Quevedo et al., 1999). 반면, 야광충이 요각류의 일일 알 생산력의 50% 이상을 섭식함에도 요각류 개체군이 영향을 받지 않은 결과도 있었다(Daan, 1987). 이와 유사한 사례로서 홍콩 폐쇄성 만에서, 요각류 알에 대한 야광충의 높은 포식압에도 불구하고 야광충 대발생 이후 요각류가 여전히 우점하였음을 보고한 바가 있다(Zhang et al., 2017). 이는 아마도 야광충이 요각류 알을 소화하는데 오랜시간이 소요되기 때문인 것과 (Daan, 1987), 섭식된 비율이 매우 낮기(Drits et al., 2013) 때문인 것으로 판단되며, 이 때문에 포식압의 영향이 예상보다 낮았던 것으로 생각되었다.
요약하면, 본 연구기간 중 출현한 야광충 식포 내에서 먹이생물이 평균 32% (24~50%)가 출현했고, 먹이생물 조성은 분변립, 원생생물, 꽃가루, 규조류, 와편모조류, 요각류, 어란, 그리고 치자어로서 매우 다양하였다. 식포 내에서 관찰된 먹이생물의 출현율은 분변립이 가장 높았고(43%), 원생생물(19%)과 꽃가루가 18%를 차지한 반면, 규조류와 와편모조류가 각각 7%와 4%를 나타내 기존연구 결과와 매우 상이하였다. 그러나 출현 시기의 수온과 먹이농도(엽록소-a) 그리고 야광충 식포 내의 먹이생물 출현율은 연구기간동안 관찰된 높은 개체수의 야광충 출현과 어느 정도 관련이 있을 것으로 생각되었다. 본 연구기간동안 야광충 식포 내의 출현율을 근거로 분변립, 원생생물, 꽃가루, 어란, 규조류, 와편모조류가 먹이로서 선호된 것으로 여겨졌다.
