

1. 서 론
2. 재료 및 방법
2.1 배양
2.2 DNA 추출
2.3 Pfiesteria piscicida 종 특이적 프라이머 제작
2.4 EvaGreen 기반 정량적 real-time PCR 기법
2.5 Pfiesteria piscicida의 한국 근해에서의 분포와 개체군 변동
3. 결 과
3.1 Pfiesteria piscicida 종 특이적 프라이머
3.2 EvaGreen 기반 정량적 real-time PCR 기법
3.3 Pfiesteria piscicida의 한국 근해 분포 및 시화호 인근 개체군 변동
4. 고 찰
1. 서 론
Pfiesteria piscicida는 Peridinales에 속하는 종속영양 와편모조류로 활성독소(TOX-A) 및 비활성 잠재독소(TOX-B)를 갖고 있다고 보고되었다(Burkholder et al., 2001). 이러한 독소는 어류에게는 피부 병변과 아가미 손상, 및 폐사를 일으키고 인간에게는 피부 병변과 호흡기 질환, 신경 질환, 단기기억상실을 유발하는 등(Burkholder et al., 2001, 2005; Burkholder and Glasgow, 1997; Grattan et al., 1998; Marshall et al., 2000; Saito et al., 2002), 환경 내 먹이사슬뿐 아니라 인간 건강에 유해한 영향을 미친다. 유해조류 P. piscicida는 1991년에 어류 폐사가 발생한 미국 북캐롤라이나의 뉴우스강과 팜리코 하구에서 최초로 발견되었으며(Burkholder et al., 1992), 미국 해안에서 발생한 다양한 어류 폐사 사건들이 P. piscicida와 연관이 있는 것으로 알려졌다(Lewitus et al., 1995; Burkholder and Glasgow, 1997). 한국에서는 이들 종이 서해와 남해안에서 분포한다고 알려졌으나(Jeong et al., 2006), 현재까지 P. piscicida로 인한 피해는 보고된 바가 없다.
세포 크기가 작은 P. piscicida는 Luciella masanensis, Cryptoperidiniopsis brodyi 등의 Pfiesteria-like dinoflagellate (PLD)및 Karlodinium micrum, K. veneficum 등과 형태적으로 유사할 뿐 아니라, 다양한 생활단계(life stage)에 따른 세포 크기(5-15 µm) 및 형태 변화(Burkholder and Glasgow, 1997; Litaker et al., 2002)로 인해, 광학 현미경만으로 현장에서 정확한 탐지 및 정량분석을 수행하는 것이 어렵다(Bower et al., 2000; Steidinger et al., 2001). 이러한 한계점을 극복하기 위해 real-time polymerase chain reaction (PCR), fluorescent in situ hybridization (FISH), sandwich hybridization, heteroduplex mobility assay, fluorescent fragment PCR 등의 다양한 분자적 분석 방법들이 개발되어왔다(Bower et al., 2000; Haywood et al., 2007; Oldach et al., 2000; Park et al., 2012, 2014; Saito et al., 2002). 이중에서 정량적 real-time PCR 기법은 대상종을 신속, 정확한 정량이 가능할 뿐 아니라 편의성 때문에, 다수의 유해조류를 대상으로 지속적으로 개발되어왔다(Bower et al., 2000; Gray et al., 2003; Galluzzi et al., 2004; Coyne et al., 2005; Park et al., 2007; Park et al., 2012, 2014).
정량적 real-Time PCR 기법은 대표적으로 형광물질이 결합된 probe를 사용하는 방법과 DNA 이중 나선구조에 결합 가능한 형광물질을 사용하는 방법이 있다. 먼저, 형광 탐침인 TaqMan probe 방법은 매우 정확하게 정량이 가능하다는 장점으로 많은 유해조류를 대상으로 활발하게 개발되어 왔으나, 경제성이 낮다는 단점이 존재한다(Bustin, 2000, 2002). 반면, DNA 삽입 형광물질을 사용하는 방법은 경제성이 상대적으로 높기 때문에, 정량적 real-time PCR 기법에 널리 사용되어 왔다(Mao et al., 2007). 그러나 대표적 DNA 삽입 형광물질인 SYBR Green은 PCR 반응을 저해, mispriming을 촉진, 프라이머-이량체(dimer)와 비특이적인 증폭산물(amplicon)과 같은 오탐(false-positive)현상 등 검출 특이성 및 민감도를 낮추는 단점(Arikawa et al., 2008)으로 인해 real-time PCR기법에는 상대적으로 적게 적용되었다. 그러나 최근 출시된 EvaGreen은 새로운 DNA 결합 형광물질로서 짧은 이중나선구조에 대한 낮은 친화성으로 PCR 저해반응을 현저하게 감소시킨 바 있다. 이와 더불어, 형광산물량의 높은 안정성, AT-rich와 GC-rich 구분 없이 DNA 이중가닥 사이에 삽입이 가능한 높은 친화력 때문에 균등하게 결합이 가능하다. 따라서, EvaGreen은 SYBR Green의 단점을 보완했을 뿐 아니라, 경제성 또한 높다는 장점을 가지고 있다(Mao et al., 2007; Park et al., 2012).
본 연구에서는 유해조류 P. piscicida에 대해 신속, 정확하고, 경제성까지 갖는 EvaGreen을 기반으로 하는 정량적 Real-Time PCR 기법을 개발하고, 한국 근해에서의 분포와 시화호에서 개체군 변동조사를 통해 현장에서의 유용성을 검증하였다.
2. 재료 및 방법
2.1 배양
실험에 사용한 미세조류는 CCMP (National Center for Marine Algae and Microbiota), NIES (Ntional Institute for Environmental Studies), NFRDI (National Fisheries Research and Development Institute), 한양대학교 등에서 분주받았으며, f/2-Si 배지(32 psu)에서 20°C, 12:12-h 명암 조건하에서 배양하였다(Table 1). 혼합영양생물인 P. piscicida (CCMP 1830, CCMP 1831)와 Pfiesteria-like dinoflagellate (PLD)에 속하는 Cryptoperidiniopsis brodyi (CCMP 2781, CCMP 2782), Luciella masanensis (CCMP 1835, CCMP 1873), 그리고 Pseudopfiesteria shumwayae (CCMP 2089, CCMP 2807)는 f/2-Si배지 (17 psu)에서 20°C, 12:12-h 명암 조건하에서, Rhodomonas sp. (CCMP 768)를 먹이원으로 제공하여 배양하였다.
Table 1. The specificity of the primer set used in qPCR with EvaGreen
2.2 DNA 추출
미세조류의 DNA를 추출하기 위해, EX 방법(Harder et al., 2003)을 사용하였다. 시료를 3.0 µm 필터(47 mm, polycarbonate filter, MILLIPORE, Ireland)에 여과시킨 후, 필터를 800 µL의 추출완충액(100 mM Tris-HCl, 100 mM Na2-EDTA, 100 mM sodium phosphate, 1.5 M NaCl, 1% CTAB)이 분주된 2 mL 마이크로 튜브(Axyzen, USA)에 옮겨 담았다. 액체질소와 water bath (65°C)를 통해 동결 및 해동을 3회 반복 후에 10 mg mL-1 농도의 proteinase K를 8 µL (Sigma, USA) 가입하여, 37°C에서 30분 처리 후, 80 µL의 20% sodium dodecyl sulfate (SDS)를 가입하고, 65°C에서 2시간 동안 반응시켰다. 이후, 888 µL의 24:1 (vol/vol) chloroform-isoamyalcohol (Sigma, USA)을 넣고, 10,000×g에서 5분 동안 원심분리 하였다. 새로운 마이크로 튜브(2 mL)에 상층액을 옮긴 뒤, 동량의 chloroform-isoamyalcohol (24:1, vol/vol)을 넣어 전과 동일한 조건에서 원심분리 후 상층액은 다시 새로운 마이크로 튜브로 옮겨 88.8 µL의 3 M sodium acetate 및 532.8 µL의 isopropanol (또는 2-propanol)과 반응시켰다. 이를 20분 동안 원심분리(14,000×g) 하고, 상층액을 조심스럽게 제거한 뒤, 1 mL의 냉각 70 % 에탄올을 넣었다. 마지막으로 14,000×g, 4°C에서 15분 동안 원심분리 하여 상층액은 조심스럽게 제거하고, 마이크로 튜브 내에 남은 용액을 공기 중에서 완전히 건조하였다. 이렇게 얻은 pellet은 100 µL 의 1× TE buffer (10 mM Tris-HCl, 1 mM EDTA; pH8)에서 용해한 후, -20°C 냉동고에서 보관하였다.
2.3 Pfiesteria piscicida 종 특이적 프라이머 제작
Pfiesteria piscicida의 종 특이적 영역을 찾기 위해, GenBank (www.ncbi.nih.gov)와 Clustal W (Thompson et al., 1997)를 이용하였다. 특이적 프라이머는 리보솜 RNA 유전자 영역 중 internal transcribed spacer 1(ITS 1) 영역을 대상으로 제작하였으며, 증폭영역의 길이는 241 bp로 제작하였다(Table 2, Fig. 2). 제작한 프라이머의 종 특이성 검증을 위해, BLAST 검색(www.ncbi.nlm.nih.gov/BLAST)을 이용하여 일차적으로 검증하였다.
Table 2. Sequence information of Pfiesteria piscicida specific primer set
| Target Species | Region | Primer | Sequence (5'-3') | Melting temperature (TM) |
| Pfiesteria piscicida | ITS 1 | PPSF2 | CCT AAT CTC ATC ACC GTG AAT | 50.5 |
| PPSR1 | GAA AGT TGT AAG AAG GAA AAT | 44.9 |
제작한 프라이머가 종 특이적 증폭이 가능한지 확인하기 위하여 대상종인 P. piscicida와 근연종 및 주요 미세조류 17종 28개 배양주(Table 1)의 PCR 실험을 수행하였다. PCR 반응은 DNA와 특이적 프라이머(0.3 µM), 10× Ex Taq buffer, 2.5 mM dNTP, 0.2 units의 Taq polymerase (Ex Taq, Takara, Japan) 최종농도로 하였다. PCR 분석의 반응조건은 initial 94°C에서 4분, main cycle 40회 반복으로 94°C에서 20초, 55°C에서 30초, 72°C에서 50초씩 반응시키고, 최종적으로 72°C에서 5분 동안 추가반응 시켰다. PCR로 증폭된 증폭산물은 1.2% 아가로스 겔(agarose gel)에서 전기영동한 후, Ethidum Bromide (Et-Br)로 염색하여 자외선 하에서 증폭산물을 확인하였다.
2.4 EvaGreen 기반 정량적 real-time PCR 기법
EvaGreen 기반 정량적 real-time PCR 기법을 위한 반응액은 1×SsoFast™ EvaGreen®Supermix (Bio-Rad, USA), 0.3 µM의 프라이머를 최종 농도로 하였으며, 반응조건은 98°C에서 3분, 40회 반복으로 98°C에서 10초, 55.5°C에서 15초 조건으로 증폭하였다. 융해곡선(melting curve)은 65 - 95°C 사이에서 관찰 되었으며, 각 단계에서 0.2°C (10초)씩 증가시키며 형광 값을 관찰하였다. 표준곡선(standard curve)을 작성하기 위해, 200배 광학 현미경하에서(Carl Zeiss, Axioplan, Germany) 계수된 4800개의 P. piscicida 세포를 앞서 언급한 EX 방법을 이용하여, DNA를 추출 후 1/10, 1/102, 1/03, 1/104배 연속희석(serial dilution)하였다. Real-time PCR 값에 대한 표준 곡선 값은 3회 반복의 연속희석된 각 시료의 CT 평균값을 이용하여, Opticon Monitor (ver. 3.0, Bio-Rad, USA) 프로그램을 통해, 직선상의 표준곡선과 계산식을 산출하였다. 또한, real-time PCR법에 의한 증폭효율성(E)은 E = 10(-1/기울기) -1 식을 이용하여 산출하였다(Revricov and Trofimov, 2006).
2.5 Pfiesteria piscicida의 한국 근해에서의 분포와 개체군 변동
2.5.1 현장 시료 채집
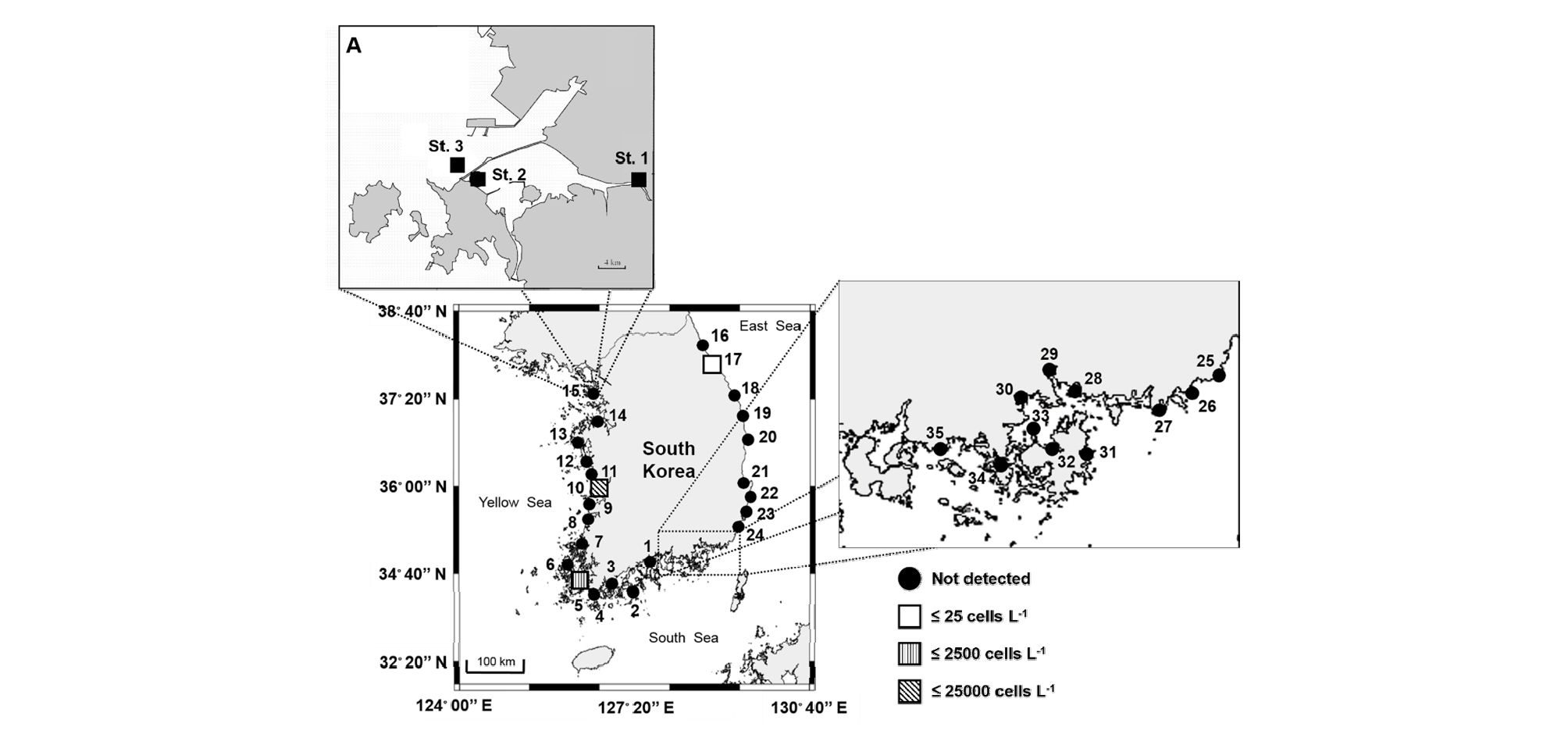
Pfiesteria piscicida의 한국 근해에서의 분포를 분석하기 위해 동, 서, 남해에서 총 35개의 정점을 선정하여, 2008년 7월부터 8월까지 표층수를 4 L씩 채수하였다(Fig. 1). 또한, P. piscicida 종의 개체군 변동을 분석하기 위해 시화호 주변 기수유역 3개의 정점(St. 1; 시화호 상류, St. 2; 배수갑문 내측, St. 3; 배수갑문 외측)에서 2007년 5월부터 2008년 1월까지 표층수를 4 L씩 채수하였다(Fig.1(a)). 본 연구의 대상 생물인 P. piscicida (5 - 15 µm)의 세포크기가 3 µm 보다 크기 때문에, 200 mL에서 1 L사이의 시료를 3.0 µm 필터(MILLIPORE, Ireland)로 현장에서 여과하여, 현장수 내 P. piscicida를 확보하였다. 필터는 DNA의 파괴를 막기 위해 800 µL의 추출 완충액(100 mM Tris–HCl, 100 mM Na2-EDTA, 100 mM sodium phosphate, 1.5 M NaCl, 1% CTAB)이 담긴 2 mL 마이크로 튜브에서 DNA 추출 전까지 -80°C 초저온냉동고(deep freezer)에서 보관하였다. 이 후, 연구실 내에서 앞서 언급한 EX 방법으로 DNA를 추출하였다. 현장 시료 내 PCR 억제자로 인한 효과를 최대한 제거하기 위해, 현장수에 우리나라에서 분포하지 않는 것으로 알려진 Chattonella subsalsa를 접종시킨 후, DNA를 추출하였다. 추출한 DNA 시료는 0, 1/10, 1/20, 1/50, 1/100로 연속희석한 후, PCR 억제효과가 나타나지 않는 최적 희석 배율을 선정하였다(Park et al. 2014).
2.5.2 물리 · 화학적 요인 분석
본 연구에서 수행한 모든 현장에서의 수온과 염분도는 수질측정 장비(Water checker U-10, HORIBA, Japan)를 이용하여 측정하였다. 영양염은 채수해온 표층수를 GF/F필터(47 mm, Whatman, USA)로 여과 후, 여과액으로 분석하였다. 영양염 분석은 각 시료마다 1회 측정되었으며, 암모니아성 질소(NH4-N)는 암모니아가 차염소산나트륨(Sodium hypochlorite) 존재 하에서 페놀(Phenol)과 반응하여 생기는 인도페놀 청색의 흡광도를 측정하여 정량하는 인도페놀법(indophenol blue method)을 이용하였고(Grkstatten et al., 1978), 아질산과 질산(NO2+NO3-N)성 질소는 하이드라진 환원 방법(Hydrazine Reduction method)으로 측정하였다(Kamphake et al., 1967). 인산염인(PO4-P)은 인산이온이 몰리브덴 산(Molibdate solution)과 반응하여 생기는 황색의 몰리브덴산착제를 아스코르빅산(Ascorbic acid)으로 환원시켜 그 흡광도를 측정하여 정량하는 아스코르빅 방법을 이용하여 분석하였다(Lowry and Lopez, 1945). 규산염은 몰리브덴산법을 이용하여 측정하였다(Bartlett, 1958). 영양염 농도와 P. piscicida와의 상관관계를 파악하기 위하여, SPSS 프로그램(ver. 23 SPSS, IL, USA )을 이용하여, Pearson 상관계수를 계산하였다.
클로로필 a는 현장수를 GF/F필터(47mm, Whatman, USA)를 이용해 일정량을 여과한 후, 필터를 90% 아세톤에서 냉암조건으로 24시간동안 추출한 후, UV-VIS spectrophotometer (Hewlett Packard, USA)를 이용하여, 각 파장별(630, 647, 664, 750 nm)로 흡광도를 측정하고, Jeffrey and Humphrey (1975)의 식에 따라 클로로필 a 값을 계산하였다.
2.5.3 생물학적 요인 분석
현장에서의 P. piscicida의 정성 및 정량 분석을 위해, 현장 시료를 추출한 DNA를 1/50 – 1/10 사이로 희석한 후, 위에서 언급한 real-time PCR 반응 조건으로 수행하였다. 실제 수계 내에는 다양한 염기서열의 DNA가 존재하기 때문에, 대상 미세조류가 아닌 것이 증폭되었을 가능성이 존재한다. 따라서 일부 현장 시료의 증폭된 PCR 산물의 염기서열을 분석한 결과, 대상 생물인 P. piscicida와 일치하는 것을 확인하였다.
모든 현장 시료는 최종 농도 1%의 무기 요오드인 루골 용액(Sigma, USA)으로 고정하였으며, 고정된 현장 시료는 조심스럽게 섞은 후 슬라이드 글라스와 400배, 1000배 배율의 광학현미경(Carl Zeiss, Axioplan, Germany)을 사용하여 식물프랑크톤을 동정하였다. 종속영양 미소 편모류의 생물량은 수단블랙(Sudan Black)용액에 2 µm 여과지(Millipore)를 염색한 후 고정된 시료를 일정량을 취해 Primulin 염색 후 1000배 배율의 광학현미경 하에서 계수하였다(Caron, 1983).
3. 결 과
3.1 Pfiesteria piscicida 종 특이적 프라이머
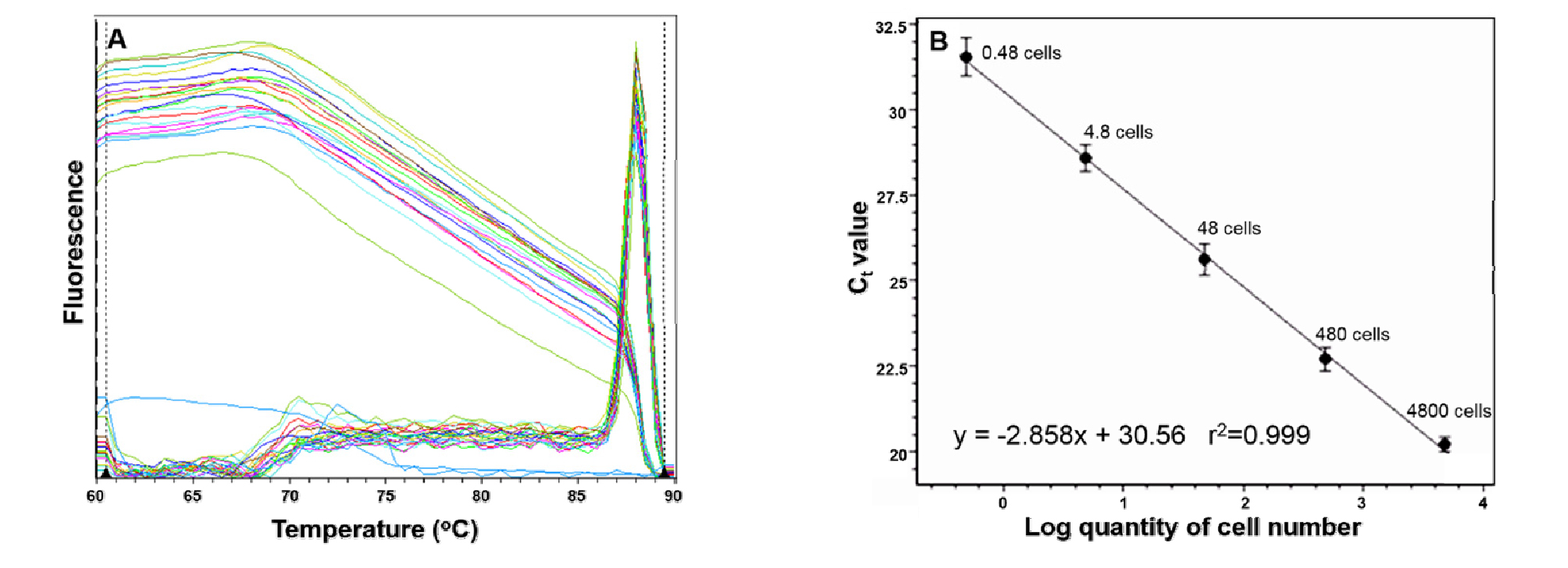
Pfiesteria piscicida 종 특이적 프라이머는 ITS 1영역을 대상으로 제작하였으며(Table 2), 이를 이용하여 P. piscicida의 DNA를 증폭 시킨 결과, 증폭산물의 길이는 241 bp로 성공적으로 증폭 되었다(Fig. 2). 제작된 프라이머의 종 특이성을 검증하기 위해, 대상종과 유전적으로 근연관계에 있는 미세조류 17종 28개 배양주(돌말류 1 종 1 배양주, 와편모조류 10 종 13 배양주, 침편모조류 6 종 10 배양주)를 대상으로 PCR 수행 결과, 대상종인 P. piscicida DNA 이외에는 증폭되지 않은 것으로 나타났다(Table 1). 이 프라이머를 이용한 EvaGreen 기반의 정량적 real-time PCR 결과, 88°C에서 하나의 피크를 나타냈을 뿐 아니라, 모드가 좁은 온도 범위(±1.3°C)를 가짐으로써, 제작된 프라이머가 P. piscicida의 대상영역만을 증폭시켰음을 알 수 있었다(Fig. 3(a)).

3.2 EvaGreen 기반 정량적 real-time PCR 기법
현장 시료내 Pfiesteria piscicida의 세포수 정량분석을 위해, 4800개의 P. piscicida 세포로부터 추출된 DNA를 1/10, 1/102, 1/03, 1/104로 연속희석 후, real-time PCR로 얻어진 각 시료들의 CT 평균값(n=3)으로 직선상의 표준곡선을 도출하였다. 그 결과, 표준곡선의 상관계수(r2≥0.999)는 매우 높게 측정되었으며(Fig. 3(b)), 증폭 효율성은 123%로 나타났다.
3.3 Pfiesteria piscicida의 한국 근해 분포 및 시화호 인근 개체군 변동
3.3.1 Pfiesteria piscicida의 한국 근해 지리적 분포
2008년 7월부터 8월까지 한국 근해 동, 서, 남해 총 35개의 정점에서 현미경 관찰로는 P. piscicida가 탐지되지 않았으나, 정량적 real-time PCR 기법을 통해 P. piscicida가 서해(정점 #5, 1,209 cells L-1; 정점 #10, 21,840 cells L-1)와 동해(정점 #10, 3 cells L-1)에 분포하고 있음을 확인하였다(Fig. 1).
3.3.2 시화호에서의 Pfiesteria piscicida의 개체군 동태
3.3.2.1 시화호 물리 · 화학적 요인 분석
시화호 정점 3개의 수온 범위는 St. 1에서 0.9 - 29.8°C, St. 2에서 1.0-31.0°C, St. 3에서는 3.4-29.0°C로 조사되었으며, 2007년 10월 이후, 모든 정점에서 수온이 감소하였다(Fig. 4(a)). 염분도는 St. 2에서 23.0-34.0 psu, St. 3에서 29.0-35.3 psu로 나타나 조사기간 동안 상대적으로 높게 유지되었던 반면, St. 1의 경우, 1.1-30.3 psu로 다른 정점에 비해 변동폭이 크게 나타났으며 특히 2007년 7월과 9월, 2008년 1월에 5 psu 이하로 낮게 나타났다(Fig. 4(b)).
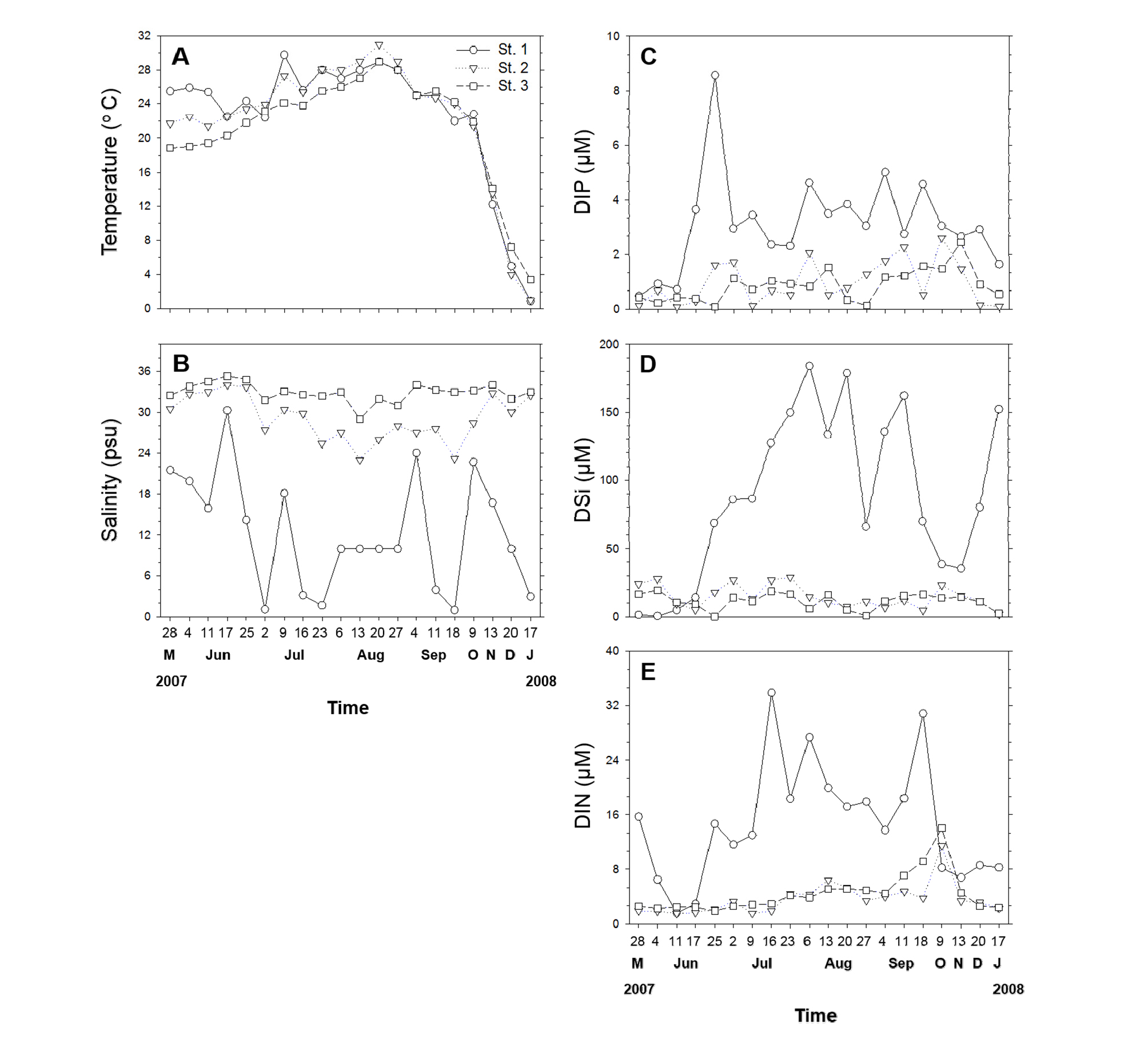
Fig. 4.
Fig. 4. Water temperature (A), salinity (B) and dissolved inorganic nutrients (DIP, dissolved inorganic phosphorus; DSi, dissolved inorganic silica; DIN, dissolved inorganic nitrogen) at three sites where located in brackish lake Shihwa from May 2007 to January 2008. Figure 4 is modified using the data in Baek et al. (2010).
용존무기규소(DISi: dissolved inorganic silicate), 용존무기질소(DIN: dissolved inorganic nitrogen), 용존무기인(DIP: dissolved inorganic phosphorus)의 영양염류는 2007년 5월을 제외한 조사기간 동안 St. 1이 다른 정점에 비해 상대적으로 높은 농도로 관찰되었다(Figs. 4(c-e)). 영양염 농도와 P. piscicida와의 상관 분석 결과, 유의한 상관관계는 관찰되지 않았다.
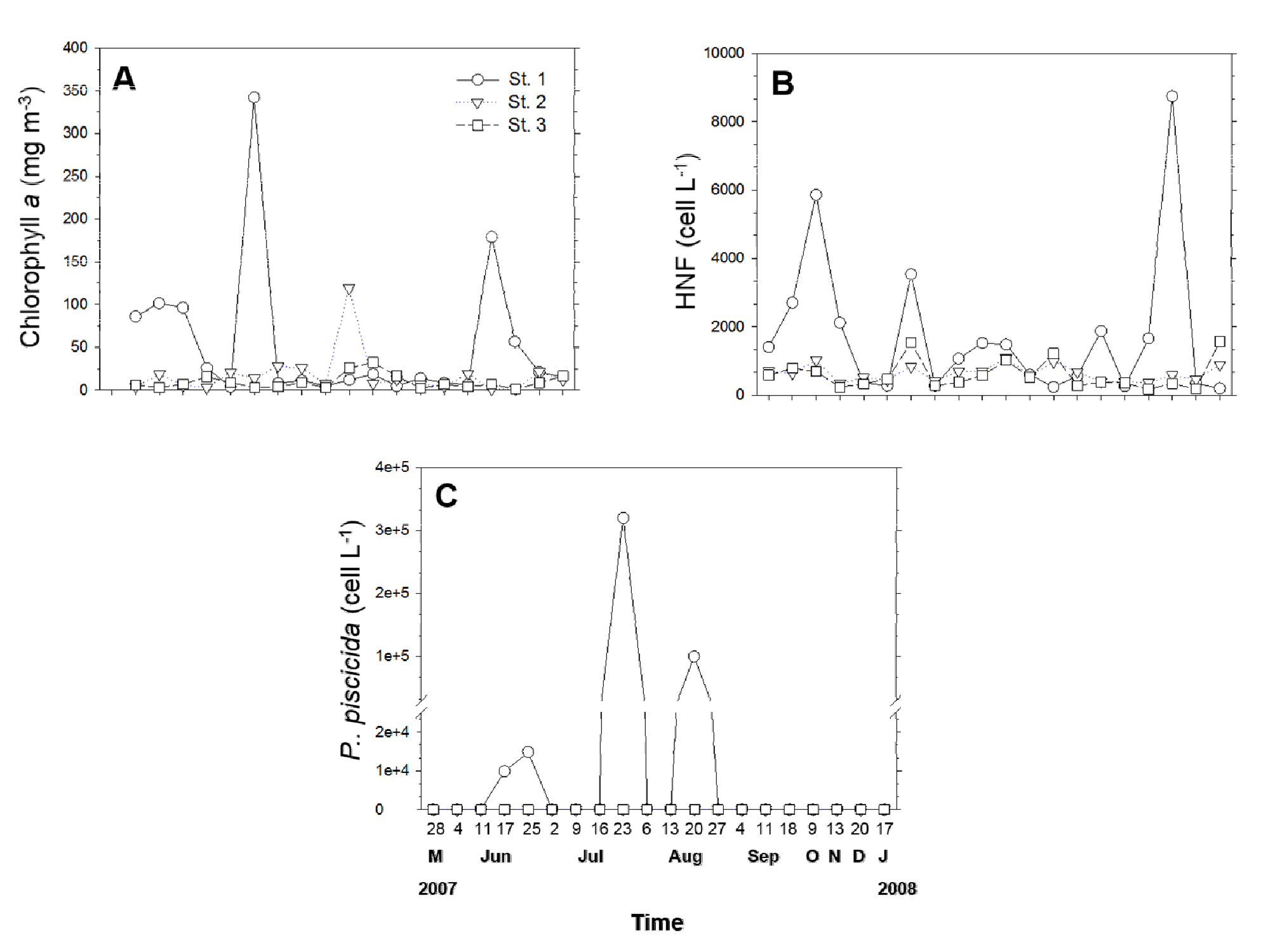
조사기간 동안 시화호에서의 클로로필 a 농도는 St. 2 (17.0 µg L-1), St. 3 (9.3 µg L-1)에 비해 St. 1 (58.5 µg L-1)에서 상대적으로 높게 측정되었다(Fig. 5(a)). St. 1의 경우, 7월에 가장 높은 클로로필 a 나타났던 반면, St. 2와 St. 3에서는 8월에 가장 높은 클로로필 a 농도가 관찰되었다.
3.3.2.2. 생물학적 요인 분석
정량적 real-time PCR 분석 결과, St. 2와 St. 3에서는 P. piscicida가 검출되지 않은 반면, St. 1에서는 2007년 6, 7, 8월에 급격한 세포수 증가가 관찰되었으며, 이때 세포 밀도 범위는 9,768-316,000 cells L-1로 나타났다(Fig. 5(c)).
P. piscicida의 먹이원으로 알려진 종속영양 미소 편모류의 평균 밀도는 St. 1에서 1.8 × 106 cells L-1로, St. 2 (6.4 × 105 cells L-1) St. 3 (6.0 × 105 cells L-1)에 비해 상대적으로 높게 나타났다(Fig. 5(b)). 조사기간 중, St. 1에서 P. piscicida의 세포수가 증가했던 2007년 6월(17, 25일)과 8월(20일)에는 종속영양 미소 편모류의 밀도가 급격하게 감소하는 현상이 관찰되었다(Figs. 5(b) and (c)).
4. 고 찰
현장에 존재하는 P. piscicida를 광학현미경으로 동정하고 정량분석을 하는 것은 이들의 세포 크기가 작고, 형태학적으로 이들과 유사한 다른 와편모조류들이 존재하기 때문에 매우 어렵다(Bowers et al., 2000; Glasgow et al., 2001; Litaker et al., 2002). 이에 대한 대안으로 TaqMan probe 기반의 정량적 real-time PCR 기법이 개발되었다(Bower et al., 2000). 그러나 TaqMan probe는 경제성이 낮아 다수의 시료 분석이 필요한 유해조류의 개체군 동태 연구 측면에서, TaqMan probe 기반 real-time PCR기법이 최적인지에 대해서는 논란의 여지가 존재한다(Kreuzer et al., 1999; Park et al., 2012). 본 연구에서는 P. piscicida를 대상으로 높은 정확성과 경제성을 가지고 있는 EvaGreen 기반의 real-time PCR기법을 개발하고, 이를 이용하여 한국 근해에서의 분포와 시화호 인근에서의 개체군 동태 조사를 통해, 개발된 기법의 유용성을 현장에서 검증하고자 하였다.
정량적 real-time PCR 기법의 종 특이성과 탐지 민감도는 DNA 대상 영역과 프라이머 제작에 의해 결정된다(Park et al., 2012). 본 연구에서 P. piscicida의 종 특이적 ITS 1 프라이머을 제작하여 검증한 결과, 제작된 프라이머는 대상종인 P. piscicida에 대해 매우 높은 특이성을 지닌 것으로 나타났다(Table 1). 정량적 real-time PCR기법은 민감도가 매우 높다고 알려져 있다. Bower et al. (2000)에 의하면, 이들에 의해서 개발된 P. piscicida 대상 real-time PCR 기법은 적은 세포 수까지도 (0.6 세포 이상) 탐지가 가능한 것으로 보고되었다. 본 연구에서 개발된 EvaGreen 기반 real-time PCR 기법의 검출 가능 세포수의 최저치는 0.48 세포로 매우 높은 민감도를 가지고 있는 것으로 판단되었다(Fig. 3(b)). 연구결과에서 나타난 높은 민감도는 종 특이적인 프라이머의 대상영역인 ITS 영역이 P. piscicida 세포 내에서 104 이상의 많은 카피 수(copy number)로 존재하기 때문인 것으로 사료된다(Guay et al., 1992; Le Blancq et al., 1997).
Real-time PCR 기법 적용 결과 도출된 표준곡선의 유의성(r2≥0.999)과 오직 하나의 피크만 나타낸 융해곡선의 결과들(Fig. 3(a))은 본 연구에서 개발된 종 특이적인 프라이머를 사용하는 EvaGreen 기반 real-time PCR 기법이 P. piscicida를 정확하게 검출 및 정량분석 할 수 있음을 나타낸다.
현장 시료에서 DNA를 추출하여 real-time PCR기법으로 대상 생물을 정량 분석할 경우, 현장 내 존재하는 다수의 PCR 억제자(부식산, 점액성 다당류, 중금속 및 페놀 화합물 등)로 인해 정확한 정량 분석이 매우 어렵다(Audemard et al., 2004, 2006; Faveri et al., 2009; Vadopalas et al., 2006). 희석법은 이러한 PCR 억제자로 인한 영향을 제거할 수 있는 것으로 알려졌다(Park et al., 2012). 본 연구에서 현장수로부터 추출한 DNA를 1/50 - 1/10로 희석하였을 때 PCR 억제효과가 제거되는 것을 확인하였기 때문에, real-time PCR기법으로 분석하기 전, 모든 현장 시료를 위에 기술된 범위로 희석하여 사용하였다.
본 연구에서 개발된 EvaGreen 기반의 정량적 real-time PCR 기법을 사용하여, 총 35개의 한국 동, 서, 남해안 시료를 분석한 결과, 서해안에 위치한 목포(정점 #5, 1,209 cells L-1)와 김제(정점 #10, 21,840 cells L-1)에서 높은 밀도로 나타났다(Fig. 1). Jeong et al. (2006)에 따르면, P. piscicida는 한국 남해안(마산만, 부산 연안, 군산, 광양, 여수)과 서해안(인천, 시화호)에서의 분포가 보고된 바 있다. 이러한 결과들은 P. piscicida가 한국 서해안에서 넓은 범위로 분포하고 있음을 나타낸다. 흥미롭게도, 이전 연구에서는 P. piscicida의 분포가 보고되지 않았던 동해에서도(강릉, 정점 #17) 낮은 농도로 P. piscicida (3 cells L-1)가 검출되었다(Fig. 1). 동해안 지역은 여름철 동한해류에 의해 남해로부터 영향을 받는것으로 알려져 있으며, 실제로 1995년과 2003년 여름철 남해안에서 Cochlodinium polykrikoides 적조 발생 당시, 강릉까지 확산된 바 있다(2013년도 한국연안의 적조발생 상황 보고서, 국립수산과학원). 따라서 P. piscicida가 강릉 지역의 자생종으로 분포하는 것인지 혹은 여름철에 해류에 의해서 남해로부터 확산되어 왔는지 여부를 판단하기 위해서는 향후 추적 관찰이 요구된다.
서해안 기수유역인 시화호를 대상으로 2007년 5월부터 2008년 1월까지 P. piscicida에 대한 정량적 real-time PCR 분석 결과, St. 1에서만 P. piscicida가 출현하였으며, 6, 7, 8월 총 세 차례의 급격한 세포수 증가가 관찰되었다(Fig. 5(c)). 이 시기 물리, 화학, 생물학적 요인의 특징은 먼저 P. piscicida가 출현한 St. 1이 다른 정점에 비해 상대적으로 낮은 염분도(≤ 25psu)가 관찰되었으며, P. piscicida가 높은 밀도를 나타냈던 시기에는 더욱 낮은(≤ 15 psu)를 염분도가 관찰되었다(Fig. 4(b)). 이러한 결과는 P. piscicida가 넓은 범위의 염분도에서 성장이 가능하며, 15 psu에서 최적 성장률를 나타낸다는 기존 연구결과(Kane et al., 1998)와도 일치 한다. 흥미로운 점은 같은 시기의 다른 정점에 비해 상대적으로 낮은 염분도(20.8, 10.5 psu)를 나타냈던 한국 서해안에 위치한 목포(정점 #5)와 김제(정점 #10)에서 앞선 결과와 유사하게 높은 밀도의 P. piscicida가 검출되었다.
영양염의 경우, St. 1이 St. 2, St. 3 정점보다 인산염, 규산염, 질산염 등의 농도가 상대적으로 높게 관찰 되었으나(Figs. 4(c-e)), 통계분석(Pearson correlation) 결과, P. piscicida와 유의적인 상관관계는 나타나지 않았다. P. piscicida는 혼합 영양성 조류이기 때문에, 영양염류의 농도뿐만 아니라, 먹이 생물의 밀도 역시도 이들 종의 동태에 영향을 미쳤을 것으로 판단된다. 실제로 P. piscicida가 가장 높은 밀도로 출현했던 2007년 7월 시료의 현미경 관찰 결과, P. piscicida의 먹이생물(Jeong et al., 2007)로 알려진 Prorocentrum sp.와 Heterosigma akashiwo가 주로 관찰되었다(data not shown). 뿐만 아니라, 한국 근해에서의 지리적 분포 조사시 P. piscicida가 높은 밀로도 출현했던 목포(정점 #5, 1,209 cells L-1)와 김제(정점 #10, 21,840 cells L-1)에서는 각각 Skeletonema costatum과 동정이 되지 않은 돌말류에 의한 조류대발생이 관찰되었다(data not shown). 그러나 P. piscicida가 이들을 실제로 섭식하여 높은 밀도를 나타냈는지는 향후 연구가 필요할 것으로 사료된다. 종속영양 미소 편모류도 P. piscicida의 먹이원으로써 알려져 있다(Jeong et al., 2007). 흥미롭게도 2007년 6월(17일, 25일)과 8월(20일), P. piscicida의 밀도가 증가했을 당시, 이들의 밀도가 현저히 감소하였는데(Figs. 5(b and c)) 이는 P. piscicida가 종속영양 미소 편모류를 섭식함으로써 증식한 것으로 사료된다. 그러나 2007년 7월(23일)의 경우, P. piscicida의 밀도가 증가하였음에도 불구하고, 6, 8월에 관찰된 현상과 상반되게, 종속영양 미소 편모류의 세포수가 오히려 소폭 증가하였다. 이는 P. piscicida의 변동이 먹이생물의 양뿐만 아니라 질의 상태에 따라서도 달라질 수 있음을 의미하며 명확하게 이들의 관계를 분석하기 위해서는 향후 차세대 염기서열 분석 방법 등의 도입이 필요할 것으로 사료된다.
앞서 언급한 바와 같이, P. piscicida는 작은 세포크기와 형태변이로 인해, 광학현미경만으로 분석하는 것은 매우 어렵다. 실제로, 본 연구에서도 채집된 모든 현장시료를 현미경 관찰했음에도 불구하고, P. piscicida는 관찰되지 않았다(data now shown). 반면, 새롭게 개발된 EvaGreen 기반 정량적 real-time PCR 기법은 동일 시료를 대상으로 P. piscicida를 분석하는데 성공하였다. 본 연구 결과는 향후, P. piscicida의 분포, 이동 및 확산 등의 생물지리학적 연구에도 크게 기여할 수 있을 것으로 사료된다.
