

1. 서 론
Entomoneis Ehrenberg 1845는 Patrick and Reimer(1975), Round et al.(1990)에 의해 전체적으로 뒤틀리고 girdle view에서 panduriform 형태를 한 세포, 수많은 girdle bands, S자 모양의 keel을 따라 위치한 raphe, 하나 또는 두 개의 엽록체, biseriate 또는 multiseriate striae, junction line (basal fibulae)의 존재 등이 분류 특징으로 확실히 확립되었다. 이후 연구에 의해 striae가 uniseriate인 종, junction line이 없는 종이 발견되어 분류 특징이 일부 수정되었으며(Al-Handal et al., 2020) 현재 AlgaeBase (Guiry and Guiry, 2024)에 35종이 기록되어 있다.
분류 특징이 확실히 확립되기 이전의 과거에는 Entomoneis가 panduriform 형태를 가지고 있는 Amphiprora Ehrenberg 1843 속으로 대부분 동정되었다. 국내에서는 이전에 동정되었던 속명인 Amphiprora (현재 Entomoneis로 재명명된 종)를 포함하여 E. alata, E. angustata (= A. angustata), E. cholnokyi (= E. costata), E. gigantea (= A. gigantea), E. gigantea var. sulcata (= A. gigantea, A. sulcata), E. paludosa (= A. paludosa), E. paludosa var. hyperborea (= A. paludosa var. hyperborea), E. paludosa var. subsalina 등 총 8종이 보고되었다(NIBR, 2023).
8종 중 참고문헌을 찾을 수 있는 종은 E. alata (Lee et al., 1994; Shim, 1994), E. angustata, E. gigantea var. sulcata, E. paludosa, E. paludosa var. hyperborea (Shim, 1994) 5종뿐이다. 이후 연구로는 광학현미경 상의 E. alata (Noh et al., 2010; Lee, 2013; Kim et al., 2018), E. gigantea var. sulcata (Moon and Choi, 2003)의 출현 연구, E. paludosa (An et al., 2017)의 형태 및 분자생물학적 연구 등 소수에 불과하여 국내에서 Entomoneis 연구가 미비한 수준임을 알 수 있다. 본 연구에서는 미기록종 3종을 발굴하고 광학, 전자현미경을 통한 형태 특성과 rbcL 염기서열 분석 결과를 제공하여 추후 국내의 Entomoneis 연구에 기초자료를 제공할 것이다.
2. 재료 및 방법
남해군과 서귀포시의 조간대에서 니질 퇴적물과 작은 자갈을 채집하였다(Table 1). 니질 퇴적물은 표층으로부터 1cm 정도의 퇴적물을 긁어서 채집하였다. 채집 시료에서 세포를 단일분리 후 배양하였다. 배양 조건은 f/2+si 배지(Guillard and Ryther, 1962)에 12:12 L:D cycle, 온도 15℃, 염분 30 psu이다.
Table 1.
Sampling date & site
| Date | Site | Substrate | Latitude (N) | Longitude (E) |
| 2023. 02. 02. | Namhae-gun | Mud | 34°49’06.9” | 127°56’29.0” |
| Gravel | ||||
| 2023. 04. 22. | Seogwipo-si | Gravel | 33°16’40.40” | 126°10’9.60” |
배양 시료는 Hasle and Fryxell(1970)의 규각 세척 과정을 따라 진행하여 광학현미경과 주사전자현미경으로 관찰하였다. 광학현미경으로 1,000 배율의 BF (bright field)로 배양 시료를, DIC (differential interference contrast illumination)로 제작한 표본을 관찰하여 세포의 전체적인 형태와 junction line을 확인하였다. 주사전자현미경으로는 세포의 길이와 폭을 측정하고, 미세구조를 관찰하였다. 폭을 측정할 때는 valve view에서 세포의 중앙 부분을 측정하였다.
DNA 추출은 AccuPrep Genomic DNA Extraction kit (BIONEER, Korea)를 사용했다. 추출한 DNA는 DprbcL1, DprbcL7 (Daugbjerg and Andersen, 1997) primer을 사용하여 rbcL (large subunit gene in chloroplast) 부분을 PCR 진행하였다. PCR 조건은 94℃ 3분 30초로 시작하여 94℃ 50초, 52℃ 50초, 72℃ 1분 30초 과정을 36회 반복한 후 72℃ 15분으로 마지막 단계를 수행하였다(Thomas et al., 2016). 얻어진 염기서열과 NCBI에서 수집한 Entomoneis의 염기서열을 Clustal W (Thompson et al., 1994)로 재배열하고 RAxML 8.2 (Stamatakis, 2014)를 사용하여 ML (maximum likelihood analysis), GTR+G bootstrapping 1,000회 실시하였다. 이 과정을 진행하여 나온 결과물 중 최적의 phylogram으로 결과를 분석하였다.
3. 결 과
미기록종 3종을 발견하였으며, 2종의 rbcL 염기서열을 얻었다.
1. Entomoneis annagodheae Al-Handal & Mucko 2020 (Fig. 1A-K)
KNU_St_23073
(1) 표본번호: MABIK DI00044252
(2) 채집지: 경상남도 남해군 이동면 초음리
(3) 채집 시료: 니질 퇴적물
(4) 채집 날짜: 2023. 02. 02
세포는 valve view에서 양쪽 중앙이 살짝 오목하게 들어가 있는 선형의 lanceolate로 길이는 47-61 𝜇m, 폭은 10-12 𝜇m이다. S자 모양의 keel을 따라 raphe가 위치한다. 중앙의 raphe canal이 살짝 열려 있어 proximal raphe가 직선으로 끝나는 것을 확인할 수 있고 distal raphe 끝은 갈고리 모양으로 심하게 휘어 있다. Girdle view에서 세포는 panduriform으로 여러 개의 girdle bands를 가진다. 세포 내부 중앙에는 규각이 두껍게 발달한 fascia가 존재한다. Junction line은 세포 전체에 이어져 있지 않고 양 끝에만 짧은 모양이다. Junction line이 세포 전체에 이어져 있지 않기 때문에 basal fibulae도 내부에 양 끝에만 불규칙한 모양으로 짧고 두껍게 존재한다. 또한 세포 전체에 이어져 있지 않은 junction line 때문에 valve의 납작한 부분인 wing의 면적이 매우 넓어 세포가 납작하고 뒤틀림이 심하지 않다. Uniseriate striae로 10 𝜇m 안에 50-58개이며 촘촘하게 배열되어 있다.
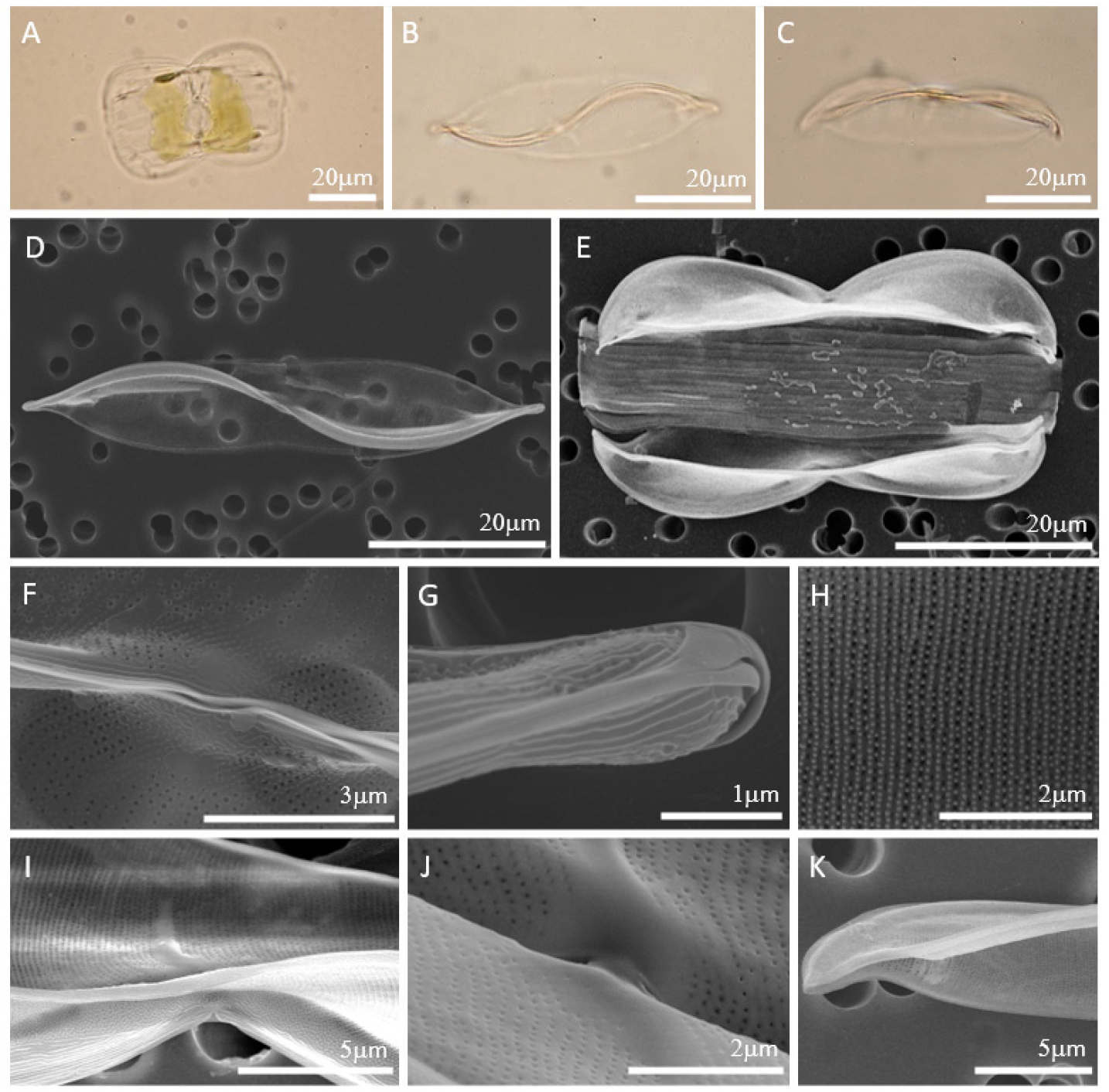
Fig. 1.
Light Microscope (LM) (Fig. 1. A-C) and Scanning Electron Microscope (SEM) (Fig. 1. D-K) images of Entomoneis annagodheae. (A) Live cell in girdle view. (B) Valve view with central fascia. (C) Girdle view with central fascia. (D) Linear lanceolate valve and slightly concave valve margins in the middle. (E) Panduriform cell with numerous girdle bands. (F) Opening central raphe canal. (G) Strongly hooked terminal raphe ending. (H) Uniseriate striae. (I) Internal valve with central fascia. (J) Details of internal central fascia. (K) Internal thickened basal fibula.
2. Entomoneis centrospinosa Osada & Kobayasi 1990 (Fig. 2A-I)
KNU_St_23090
(1) 표본번호: MABIK DI00044253
(2) 채집지: 경상남도 남해군 이동면 초음리
(3) 채집 시료: 작은 자갈
(4) 채집 날짜: 2023. 02. 02
세포는 girdle view에서 panduriform으로 여러 개의 girdle bands를 가지고 있으며 길이는 88-91 𝜇m이다. Valve의 납작한 부분인 wing은 매우 납작하며 뒤틀림이 심하다. S자 모양의 keel을 따라 raphe가 위치하며 proximal raphe 끝은 약간 확장되어 있고 distal raphe 끝은 약하게 휘어 있다. 세포 중앙의 raphe 양옆으로 0-2개의 가시가 존재하며 상각과 하각에 똑같이 존재하지 않을 수 있다. Junction line은 아치형으로 세포 전체에 이어져 있기 때문에 내부의 basal fibulae도 junction line을 따라 세포 전체에 이어져 있다. Biseriate striae로 10 𝜇m 안에 18개이다. Valve body의 costae가 wing의 costae보다 더 두껍고, areola는 선형의 hymen으로 막혀 있다.
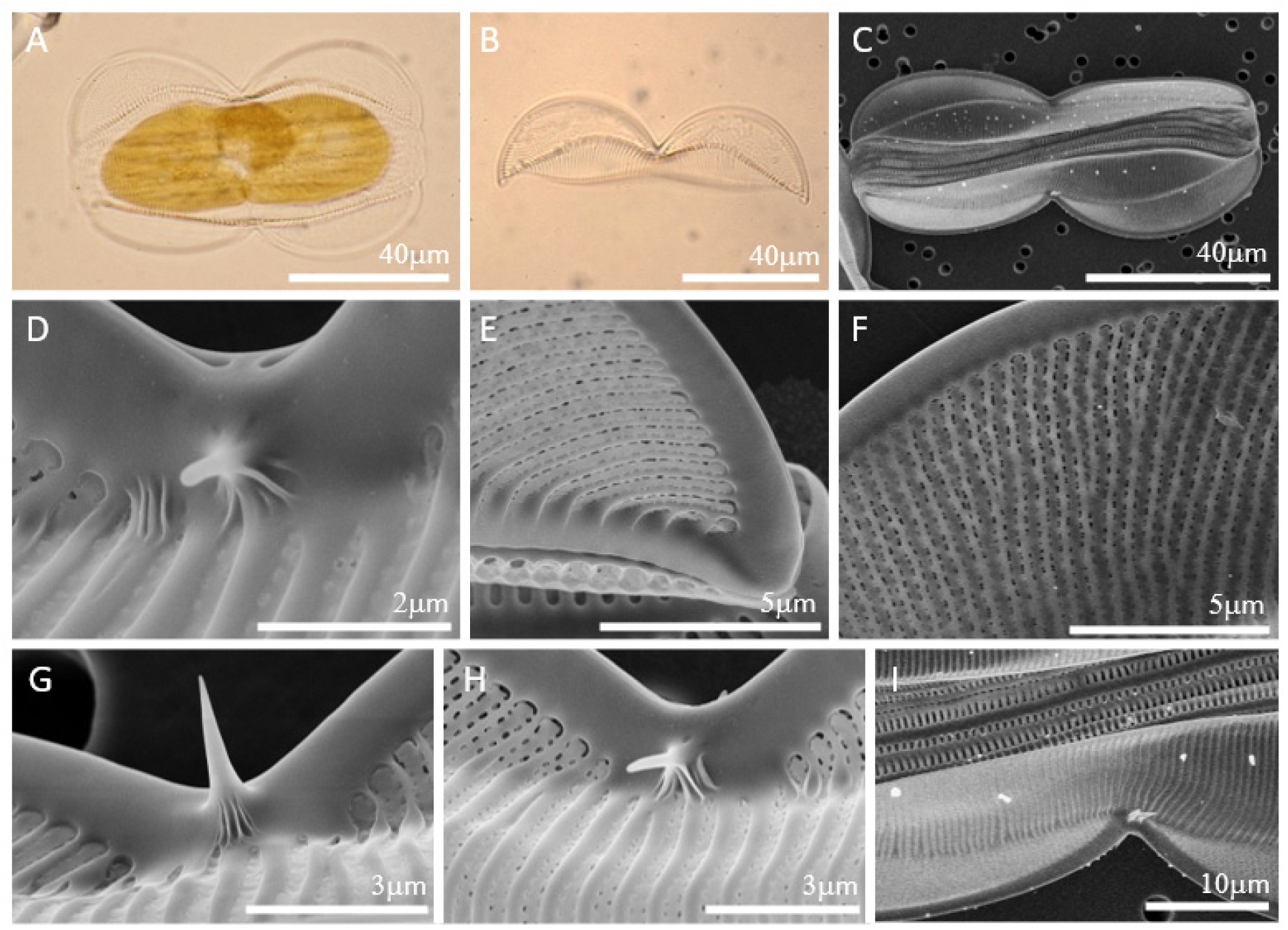
Fig. 2.
LM (Fig. 2. A-B) and SEM (Fig. 2. C-I) images of Entomoneis centrospinosa. (A) Live cell in girdle view. (B) Arcuate junction line. (C) Panduriform cell with numerous girdle bands. One valve without spine, the other valve with 2 spines. (D) Expanded proximal raphe endings. (E) Hooked terminal raphe ending. (F) Biseriate striae occluded by linear hymen. (G) One spine on the central nodule. (H) One spine on each side of the central nodule. (I) Two spines on the central nodule.
3. Entomoneis punctulata (Grunow) Osada & Kobayasi 1990 (Fig. 3A-H)
KNU_St_23068
(1) 채집지: 경상남도 남해군 이동면 초음리
(2) 채집 시료: 작은 자갈
(3) 채집 날짜: 2023. 02. 02
KNU_St_23300
(1) 표본번호: MABIK DI00044254
(2) 채집지: 제주특별자치도 서귀포시 신도포구
(3) 채집 시료: 작은 자갈
(4) 채집 날짜: 2023. 04. 22.
규각이 얇고 약해서 잘 깨지므로 세포의 온전한 형태를 확인하기 어렵다. 세포는 valve view에서 가운데 폭이 가장 넓고 양 끝으로 갈수록 얇아지는 lanceolate로 길이는 24-51 𝜇m, 폭은 9-11 𝜇m이다. S자 모양의 keel을 따라 raphe가 위치하며 proximal raphe와 distal raphe는 직선으로 끝난다. 세포는 girdle view에서 panduriform으로 여러 개의 girdle bands를 가진다. Junction line은 세포 전체에 이어져 있지 않고 양 끝에만 짧은 아치형이기 때문에 basal fibulae도 내부의 양 끝에만 짧은 아치형으로 존재한다. 매우 희미하고 약한 striae를 가져 잘 관찰되지 않는다. 희미한 striae로 인해 상대적으로 costae가 두껍게 발달한 것처럼 보인다. Hymen으로만 이루어진 biseriate striae (areola가 두 줄로 배열된 것이 아닌, 오직 hymen으로만 구성)로 10 𝜇m 안에 36-40개이다.
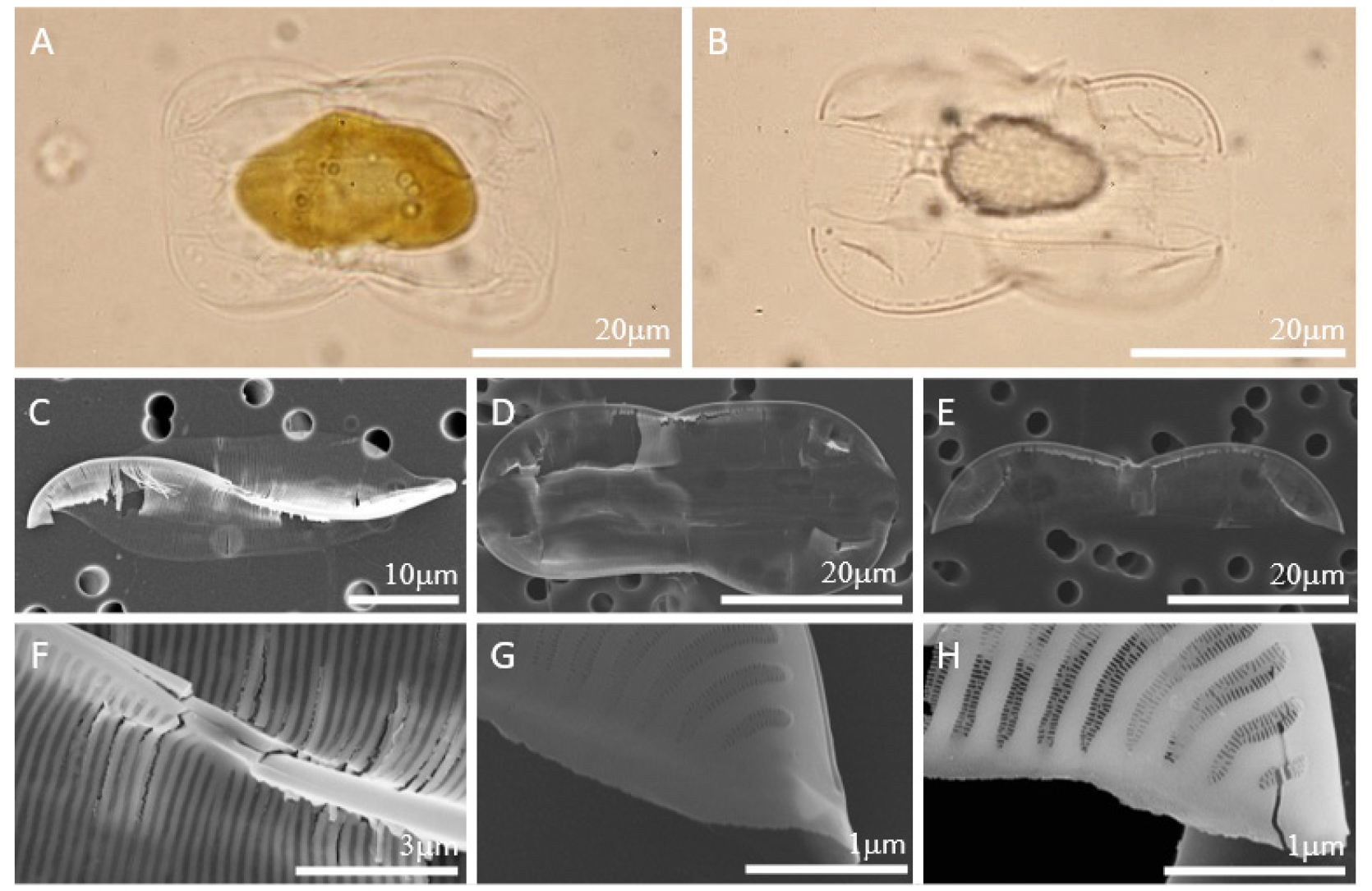
Fig. 3.
LM (Fig. 3. A-B) and SEM (Fig. 3. C-H) images of Entomoneis punctulata. (A) Live cell in girdle view. (B) Partially arcuate junction line. (C) Lanceolate weak valve with the sigmoid keel. (D) Panduriform cell with numerous girdle bands. (E) Partially arcuate basal fibulae exist along the restricted junction line. (F) Straight proximal raphe endings. (G) Straight distal raphe ending. (H) Biseriate striae composed of hymen.
4. 토 론
Entomoneis는 girdle view에서 panduriform의 형태를 가진다. 이는 Amphiprora에서 동일하게 나타나는 특징으로 과거에는 두 속을 명확하게 구분하는 분류 특징이 없어 Amphiprora 속으로 대부분 동정되었다. 이후 연구가 진행되면서 세포가 전체적으로 뒤틀리고 S자 모양의 keel과 junction line을 가진 종을 구분하면서 몇몇 종이 Amphiprora에서 Entomoneis 속으로 이동하였다. 하지만 junction line이 세포 전체에 이어지지 않고 부분적으로 존재하거나 아예 존재하지 않는 종이 발견되면서 여전히 분류 특징이 추가되고 있다. 따라서 전체적으로 뒤틀리고 girdle view에서 panduriform 형태를 가지고, S자 모양의 keel을 가진 세포를 Entomoneis로 보고 junction line의 유무와 모양, 미세구조를 확인하여 분류해야 한다.
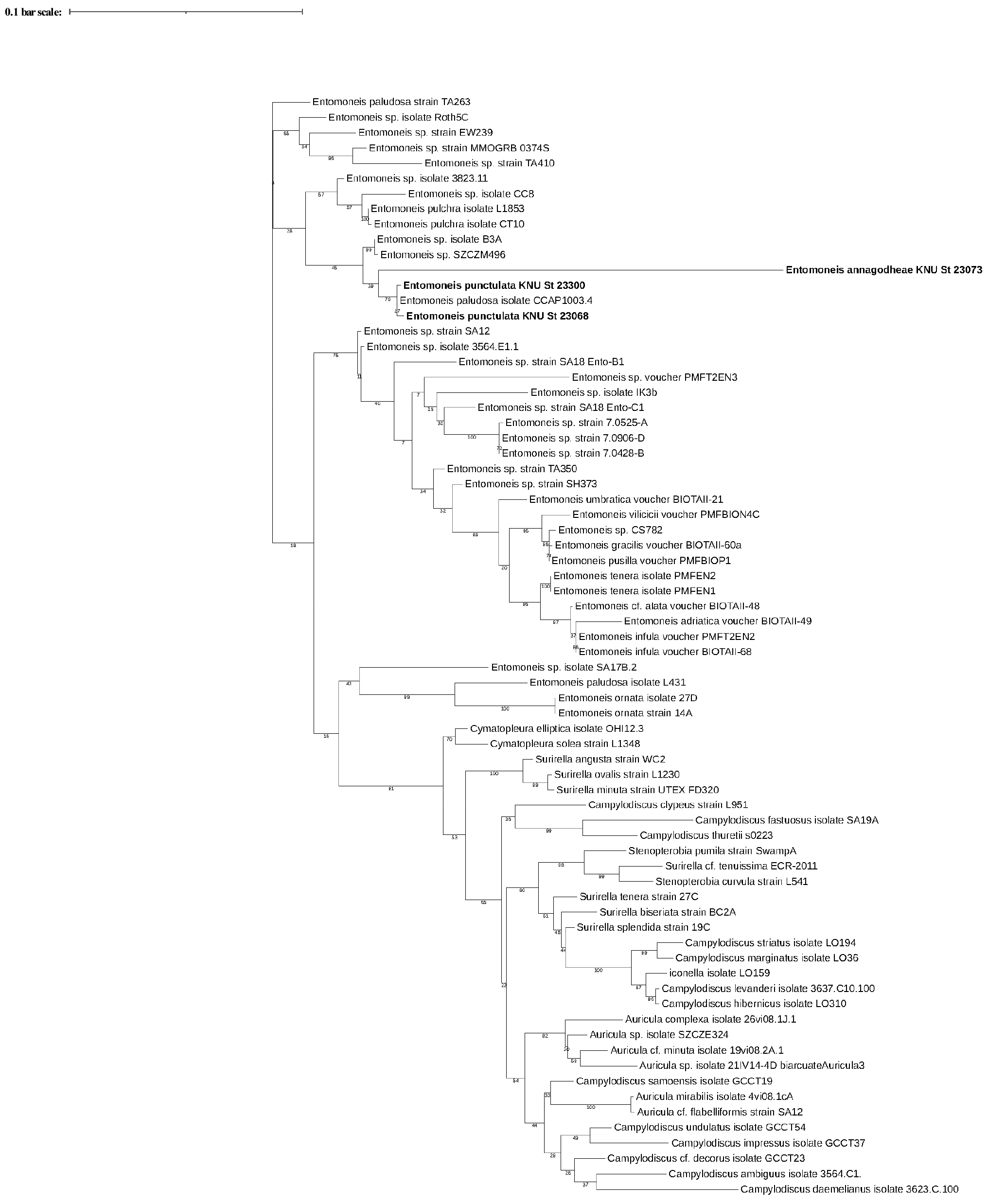
E. annagodheae는 세포 내부 중앙에 fascia가 존재한다. 이는 35종의 Entomoneis 중에서 유일하게 존재하는 독특한 특징이다. 국내에서 출현한 E. annagodheae에도 세포 내부 중앙에 fascia가 존재하였으며 Al-Handal et al.(2020)에서의 형태와 길이, 폭, 10 𝜇m 안의 striae 개수가 일치한다(Table 2). E. annagodheae는 보고된 DNA 연구 결과가 없지만 본 연구에서 rbcL 염기서열을 얻었다. 얻은 염기서열을 통해 작성한 phylogenetic tree (Fig. 4)에서 branch의 길이가 다른 종들에 비해 매우 길다. Branch의 길이가 길다는 것은 다른 종과는 구분되는 특징이 있다는 것을 의미하며 E. annagodheae는 다른 종에서 나타나지 않는 유일한 특징인 fascia를 가지기 때문에 branch가 긴 것으로 판단된다.
E. centrospinosa는 central nodule의 양쪽으로 0-3개의 가시가 존재한다. Osada and Kobayasi(1990a)는 central nodule 양쪽에서 0-3개의 가시가 존재하고, 보통 상각과 하각 중 하나의 valve에만 존재하며 central nodule 양쪽 가시의 개수가 일치할 수도 일치하지 않을 수도 있다고 설명한다. 국내에서 출현한 E. centrospinosa에서는 central nodule 양쪽으로 0-2개의 가시가 존재했다. 전자현미경으로 valve view 모습을 관찰하지 못해 폭을 측정하지 못했지만, Osada and Kobayasi(1990a)에서의 형태와 길이, 10 𝜇m 안의 striae 개수가 일치한다(Table 2).
E. punctulata는 매우 희미하고 hymen으로 이루어진 biseriate striae와 세포 전체에 이어지지 않고 양 끝에 짧은 아치형의 junction line이 나타난다. 세포 전체에 이어지지 않고 제한되어 있는 아치형의 junction line은 독특한 특징이다. 국내에서 출현한 E. punctulata도 같은 junction line을 가지며 Osada and Kobayasi(1990b)에서의 형태와 길이, 폭, 10 𝜇m 안의 striae 개수가 일치한다(Table 2). E. punctulata는 보고된 DNA 연구 결과가 없지만 본 연구에서 얻은 rbcL 염기서열을 통해 작성한 phylogenetic tree (Fig. 4)에서 Entomoneis paludosa isolate CCAP1003.4와 같은 종으로 확인되었다. 하지만 Entomoneis paludosa isolate CCAP1003.4가 발표된 Ruck et al.(2016)에서 종에 관한 설명과 사진이 없어서 E. paludosa인지 확인할 수 없었다. 또한 E. paludosa는 물결모양(sinuous)의 junction line (Tatenda et al., 2015)을 가지는 반면, E. punctulata는 세포 전체에 이어지지 않고 제한되어 있는 아치형의 junction line을 가지므로 E. paludosa와는 형태적으로 완전히 다르다.
Entomoneis는 세포가 살아있는 상태에서 분류 특징인 junction line을 관찰하기 어렵고 관찰되었다고 해도 대부분이 아치형의 junction line을 가지기 때문에 광학현미경에서 종을 구분하기 어렵다. 유사한 종 사이의 차이는 stria가 몇 줄 배열인지, areola는 어떤 hymen으로 막혔는지, 독특한 미세구조가 존재하는지 등에 따라 종이 구분되지만 이러한 특징은 광학현미경에서 관찰하기 어렵다. Entomoneis의 국내 연구에서는 광학현미경을 통한 출현 연구가 주로 이루어졌고 국내에 기록된 8종 중 E. paludosa, E. paludosa var. subsalina를 제외한 6종이 모두 아치형의 junction line을 가진다. 또한 종을 확인할 수 있는 설명과 사진이 없어 오동정이 되었을 가능성이 있어 제대로 분류되었는지 판단할 수 없다. 따라서 국내의 Entomoneis의 연구 발전을 위해서는 광학현미경에서 형태를 확인하는 것도 중요하지만, 미세구조를 확인할 수 있는 전자현미경에서의 관찰이 필요하다. 추가적으로 DNA 연구도 함께 진행되면 국내에서 다른 미기록종이나 더 나아가서 신종도 발굴할 수 있다고 사료된다.
Table 2.
Comparison of 3 species. (Width at valve view / -: no data)
| Species |
Length (𝜇m) |
Width (𝜇m) |
Striae in 10 𝜇m | Row | Junction line | Reference |
| E. annagodheae | 50-82 | 10-21 | 52-58 | 1 | Partially arcuate | Al-Handal et al., 2020 |
| E. annagodheae (KNU_St_23073) | 47-61 | 10-12 | 50-58 | 1 | Partially arcuate | This study |
| E. centrospinosa | 50-110 | 10-13 | 14-20 | 2 | Arcuate | Osada and Kobayasi, 1990a |
| E. centrospinosa (KNU_St_23090) | 88-91 | - | 18 | 2 | Arcuate | This study |
| E. punctulata | 18-53 | - | 34-36 | 2 | Partially arcuate | Osada and Kobayasi, 1990b |
| E. punctulata (KNU_St_23068, 23300) | 24-51 | 9-11 | 36-40 | 2 | Partially arcuate | This study |
