

1. 서 론
2. 재료 및 방법
2.1 대형저서동물 채집 및 저서환경 조사
3. 결 과
3.1 저서환경
3.2 종수, 서식밀도, 생물량 분포
3.3 우점종 분포
3.4 집괴분석과 정점군 특성
3.5 상관관계 및 주성분분석
4. 고 찰
4.1 대형저서동물 종 특성
4.2 주요 우점종 특성
4.3 군집구조 및 상관분석
1. 서 론
해양수산부는 지난 2006년부터 ‘해양생태계 보전 및 관리에 관한 법률’을 제정하여 우리나라 연근해를 10년 주기로 조사하도록 하였다(Kim, 2019). 2014년 이후 법을 개정하여 10년 주기는 삭제하고 ‘국가해양생태계종합조사’로 명칭을 통합하여 격년으로 조사를 실시하고 있다(Kim, 2019). 조사항목은 부유·저서환경과 부유·저서·유영생물 군집 분야이며, 이들 항목에 대한 종합적인 연구를 통해 해양생태계의 장·단기 변동 특성을 체계적으로 파악함으로서 우리 바다 해양생태계를 종합적이고 효율적으로 관리할 수 있는 기반을 마련하고자 하였다.
대형저서동물은 퇴적물과 수층의 경계면에 서식하고 한번 정착하면 이동성이 적어 군집의 종조성 변화는 해양환경 변화를 잘 반영하는 동물군이다(Snelgrove and Butman, 1994). 또한 생활사는 동물플랑크톤이나 중형저서동물에 비해 상대적으로 복잡하기 때문에 지속적인 환경변화요인이나 급격하게 변화하는 환경 영향을 잘 반영하고 있다. 이러한 특성으로 인하여 저서생태계의 장기적인 변화나 건강도를 평가하는데 중요한 지시자로 이용되고 있다 (Rakocinski et al., 2000; Reiss and Kröncke, 2005; Blanchet et al., 2008; Wildsmith et al., 2011; Peng et al., 2013).
조하대 연성저질 환경에서 대형저서동물의 분포를 결정하는 다양한 환경요인 중 일차적인 요인으로 퇴적환경을 들 수 있다. 특히 퇴적물 입도는 유속, 유기물 함량, 중금속, 유기화합물 등 다양한 환경요인들과 연관되어 있어 중요한 환경인자로 알려져 있다(Gray, 1974; Buchanan, 1984; Snelgrove and Butman, 1994). 그러나 해역별 특징으로 인해 다양한 해양환경조건이 존재하는 해역에서는 퇴적물 뿐만 아니라 저층해수 특성이나 수심이 저서동물의 분포를 결정하는 중요한 요인으로 작용할 수 있다(Choi and Koh, 1988; Hong et al., 2000; Kang et al., 2014; Seo et al., 2016). 또한 연안에 설치되는 구조물에 의한 조류 방향 변화, 연안역의 준설 등도 대형저서동물군집의 분포를 변화시킬 수 있다(Ryu et al., 1997; Sato and Azuma, 2002; Choi et al., 2003; Yu et al., 2011; Lee, 2013).
우리나라 연안역에서 수행된 저서대형동물 군집에 대한 연구는 주로 하구역, 내만해역과 도시주변에 집중되어 왔다(Shin, 1995; Shin et al., 2001; Paik et al., 2007; Lim, 2015; Kim et al., 2016; Lim et al., 2016; Yoon et al., 2017; Bae et al., 2017). 또한 동해 대륙붕 및 대륙사면에 걸치는 해역에 대한 연구(Choi and Koh, 1988, 1989, 1990), 독도 근해 심해역에 대한 연구(Choi et al., 2002), 제주도 연안해역에 대한 연구(Ko et al., 2008, 2011) 등의 수많은 저서동물군집 연구가 수행되어 왔으나 우리나라 동,서,남해를 모두 포함하는 전해역을 대상으로 동시간대에 수행한 연구는 전무한 실정이다.
본 연구의 목적은 우리나라 동,서,남해 전연안을 대상으로 전반적인 대형저서동물의 공간 분포를 파악하고 집괴분석을 통하여 저서동물 군집을 구분한 후 이들을 저층 해양환경 요소들과 연관지어 해석하고자 하였다. 본 연구결과는 한반도 연안역의 저서생태계에 대한 전반적인 이해와 효율적인 관리방안 마련을 위한 기초자료로 활용될 수 있을 것이다.
2. 재료 및 방법
2.1 대형저서동물 채집 및 저서환경 조사
우리나라 서해안의 인천부터 동해안의 속초에 이르는 한국 연안해역에서의 대형저서동물 군집 구조를 파악하기 위해 2017년 8월에 총 117개 정점을 선정하고 현장 조사를 수행하였다(Fig. 1). 각 정점에서의 대형저서동물 채집을 위해서는 표면적 0.1㎡의 van Veen grab 채집기를 이용하여 정점마다 3회씩 반복하여 해저 퇴적물을 채집하였다. 선상으로 인양된 퇴적물은 망목 1.0 mm 체에 넣고 해수를 사용하여 펄을 제거하였으며, 체에 남은 시료를 10% 중성 포르말린으로 고정하여 실험실로 운반하였다. 실험실에서 핀셋을 이용하여 저서동물을 선별하였으며, 해부현미경과 광학현미경으로 저서동물을 종수준까지 동정하였다. 종별로 개체수를 기록한 후 0.001 g까지 측정할 수 있는 저울을 사용하여 종별 습중량을 측정하였다. 군집분석용 자료는 종(열)×정점(행)으로 구성된 행렬자료(matrix)로 작성하였으며, 서식밀도와 습중량은 1 m2당 수치로 환산하여 나타내었다.
저층환경요인 중 수심은 각 정점에서 CTD (RBRconcerto)를 이용하여 측정하였고, 퇴적물 시료는 대형저서동물 채집과는 별도로 채취한 van Veen grab 내 표층퇴적물을 채취하여 분석하였다. 퇴적물의 분석항목은 평균입도, 분급도, 사질함량, 실트함량, 클레이함량, 유기물함량(Loss On Ignition), 총유기탄소량 등이었다. 각 항목의 해양환경공정시험법(MOF, 2013)에 의거하여 분석하였다.
각 조사정점에서의 대형저서동물군집의 특성을 파악하기 위하여 생태지수인 종다양도(H′), 종풍부도(R), 종균등도(J), 우점도(D)를 계산하였으며, 대형저서동물의 공간 분포를 파악하기 위하여 집괴분석을 수행하였다. 집괴분석은 모든 저서동물을 대상으로 분석을 실시하였고, 자료 편중을 피하기 위해 출현개체수를 log (x+1)로 대수변환을 하였다. 여기서 x는 각종의 출현 개체수 자료이다. 유사도 지수는 Bray-Curtis similarity (Bray and Curtis, 1957)로 계산하였고 정점군은 Group average mode 결합법을 이용하여 수지도를 그린 후 유사도 x%에서 구분하였다. 또한 SIMPER 분석을 통하여 각 정점군에 기여율이 높은 우점종들의 분석하였다. 그리고 저서군집에 영향을 주는 환경요소를 파악하기 위하여 각 환경자료에 대한 변수 값을 표준화(Normalization)한 후 Bio-Env 분석을 실시하였다. 대형저서동물군집과 저서환경요인간의 상관관계를 파악하기 위하여 Spearman rank correlation coefficient를 구하였으며, 조사해역의 저서환경요인(8개), 우점종(13종)의 분포와 생태지수(4개)간의 연관성을 알아보기 위하여 주성분분석(Principal Component Analysis; PCA)을 수행하였다. 환경자료 변환은 사질, 실트, 클레이함량은 arcsin (√x)변환 하였고, 평균입도, 분급도, 유기물함량, 총유기탄소량은 log (x+1)변환하여 분석하였다(Zar, 1984). 본 논문에서 사용한 프로그램은 Primer (ver. 6) 및 SPSS (ver. 23)를 이용하였다.
3. 결 과
3.1 저서환경
한국 연안해역의 총 117개 정점에서의 평균 수심은 24.2±15.2 m 이었으며, 서해, 남해와 동해해역으로 구분해서 보면 서해는 20.3±10.5 m, 남해 21.1±9.9 m, 동해 53.3±17.5 m로 동해가 다른해역에 비해 깊은 수심을 나타났다. 입도특성을 보면, 사질함량은 서해해역(60.0%)이 남해(26.8%)나 동해해역(32.9%)보다 높은 값을 보였으며, 실트함량은 남해해역(52.2%)이 서해(29.4%)나 동해해역(48.6%)에 비해 높은 값이 나타났다. 유기물함량(LOI)은 남해(6.9%), 동해(5.5%), 서해(3.7%)해역순으로 높은 함량을 보였다(Table 1).
Table 1.
Distribution of grain size and LOI of surface sediment, station number and Depth along each survey areas in Korean coastal waters in August 2017
조사해역별로 비교해 보면, 서해해역은 수심과 유기물함량이 낮고 사질함량이 높은 해역이고 남해해역은 서해해역과 비슷한 수심을 보이지만 실트함량과 유기물함량이 높은 해역이였다. 동해해역은 남해해역에 비해 수심이 깊으나 실트함량과 유기물함량은 비슷한 수준으로 나타났다.
3.2 종수, 서식밀도, 생물량 분포
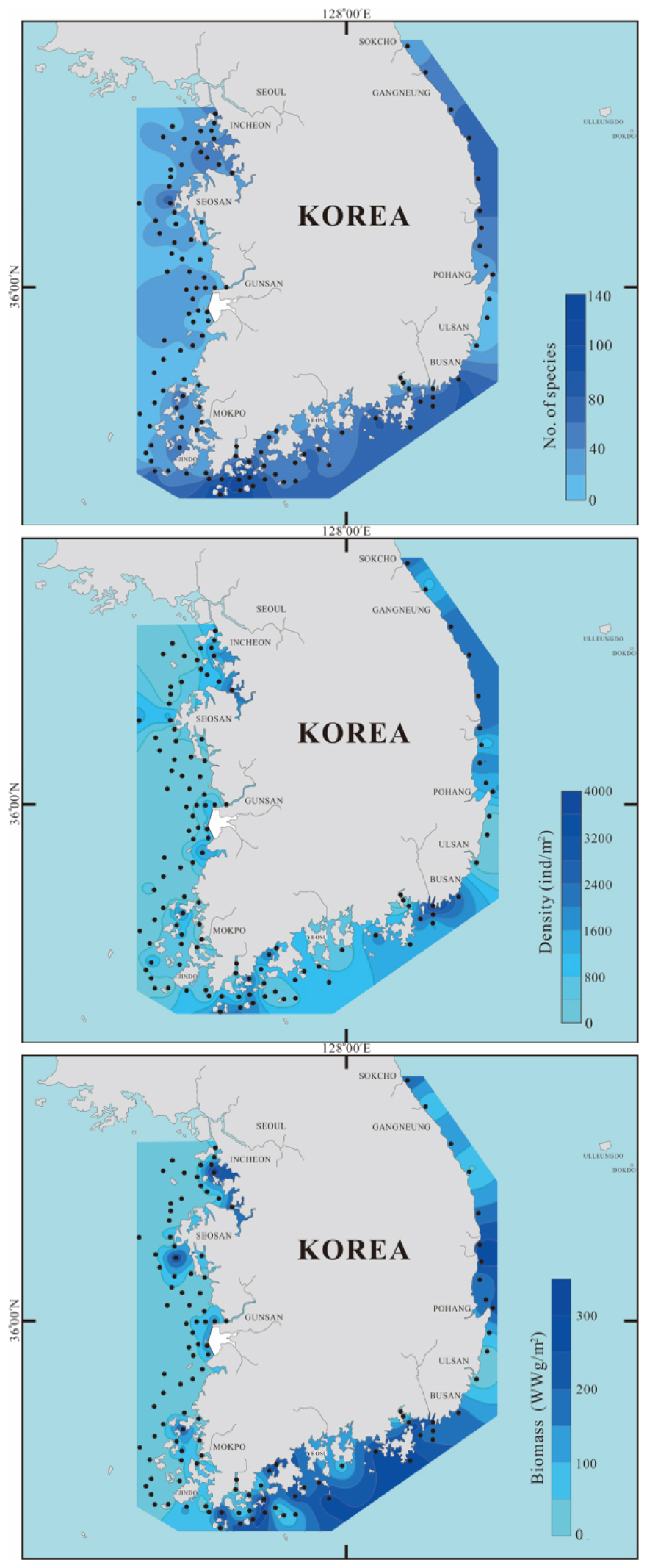
한국 연안해역의 117개 정점에서 출현한 대형저서동물은 총 613 종/35.1 m2이었다(Table 2). 환형동물이 233 종/35.1 m2으로 전체 출현종수의 38.0%를 차지하여 가장 우점하였으며, 절지동물 172 종/35.1 m2, 연체동물 155 종/35.1 m2, 극피동물 28 종/35.1 m2이 출현하였다. 그 밖에 자포동물, 유형동물, 성구동물 등 기타 동물군에서 25종/35.1 m2이 나타났다. 조사해역 전체 평균 서식밀도는 1,228 개체/m2였으며, 동물군별 서식밀도는 출현종수와 마찬가지로 환형동물(916 개체/m2), 절지동물(161 개체/m2), 연체동물(99 개체/m2), 극피동물(29 개체/m2)의 순서로 출현하였으며, 기타 동물군은 23 개체/m2의 서식밀도를 보였다. 평균 생물량(습중량)은 110.9 gWW/m2이었으며 극피동물(59.1 gWW/m2)이 전체의 53.3%를 차지하여 생체량에서 우점 분류군이었다. 그 다음으로 연체동물(33.6 gWW/m2), 환형동물(10.3 gWW/m2), 기타 동물군(4.3 gWW/m2), 절지동물(3.7 gWW/m2)의 순서로 높은 값을 보여 출현종수 및 서식밀도와는 다른 양상을 보였다.
출현종수, 서식밀도, 생물량의 공간분포 양상은 Fig. 2와 같다. 출현종수는 금강하구 입구해역 정점 W25, 함평만 주변 정점 W38과 마산만 정점 S39에서 가장 적은 2종이 출현하였다. 90종 이상 출현한 정점은 모두 완도해역 부근에 위치한 7개 정점(정점 S3, 7, 8, 9, 49, 50, 51)이었다. 출현종수가 적은 정점들은 모두 하구역 입구나 만 내부에 위치한 정점들이었다. 출현종수가 많은 정점들에서는 환형동물이 60% 이상, 절지동물이 20% 정도 차지하였다. 서식밀도는 낙동강 입구해역인 정점 S45에서 40,649 개체/m2가 출현하여 가장 높은 밀도를 보였는데, 이는 얼굴갯지렁이류인 Pseudopolydora kempi가 극우점한 것에 기인하였다. 2,000 개체/m2 이상의 밀도를 보인 정점들은 주로 완도 주변 및 부산과 동해해역에 위치하여 전체적으로 보아 서해보다는 남해 및 동해해역에서 높은 서식밀도를 보였다. 생물량은 남해와 동해해역에서 대부분의 정점에서 높은 값을 보였고, 서해는 연안 부근에서 비교적 높은 값을 보였다. 서해와 남해해역은 성게류, 동해해역은 거미불가사리류의 출현이 생물량 증가에 관련이 있는 것으로 나타났다.
Table 2.
Number of species, density and biomass of major faunal groups of macrobenthos in Korean coastal waters in August 2017
3.3 우점종 분포
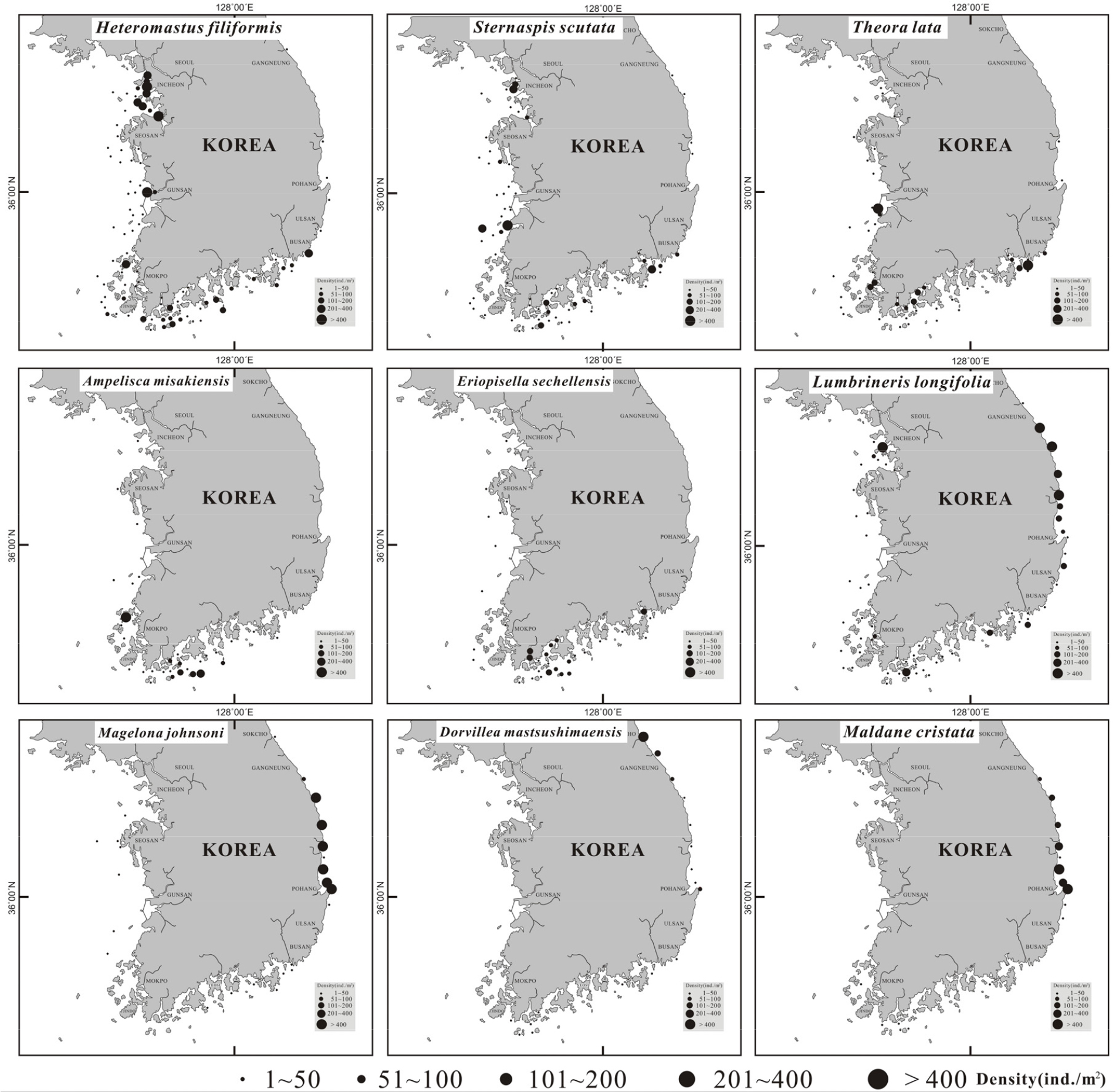
한국 연안에 분포하는 대형저서동물 가운데 상위 1% 이상을 차지하는 우점종은 총 15종 이었고, 동물군별로는 환형동물에서 11종, 절지동물에서 3종, 연체동물에서 1종이었다(Table 3). 상위 우점종 15종이 전체 밀도에서 차지하는 점유율은 약 60%였다. 이들의 공간분포 양상을 보면(Fig. 3), 첫번째와 열한번째 우점종인 얼굴갯지렁류 Pseudopolydora kempi, Rhynchospio sp.는 낙동강 하구입구 정점(S45)에서만 제한적으로 출현하였고, 각각 37,000 개체/m2와 2,200 개체/m2의 밀도로 극우점하였다. 이 종들은 출현하는 빈도가 적어서 낙동강 하구 정점을 제외하면 거의 채집되지 않았다. 두 번째 우점종인 버들갯지렁이류인 Heteromastus filiformis와 오뚜기갯지렁인 Sternaspis scutata는 동해해역을 제외하고 대부분의 정점에서 각각 3-3,540 개체/m2와 3-573 개체/m2 밀도로 출현하였으며, 정점별 서식밀도의 차이가 크게 나타났다. 세 번째 우점종인 양손갯지렁이류 Magelona jaohnsoni와 아홉 번째 우점종인 대나무갯지렁이류 Maldane cristata는 서해와 남해해역보다는 동해해역에서 수심이 비교적 깊은 해역에서 각각 3-2,924 개체/m2 및 3-669 개체/m2 출현하였다. 네 번째 우점종인 송곳갯지렁이류 Lumbrineris longifolia는 전체 정점수의 약 50% 이상인 60개 정점에서 출현하였으며 서식밀도는 3-736 개체/m2였다. 동해해역이 다른 해역보다는 높은 밀도를 보였다. 옆새우류인 Ampelisca misakiensis와 Eriopisella sechellensis는 대부분 남해 해역에 위치한 정점들에서 출현하였으며, 각각 3-436 개체/m2 및 3-160 개체/m2 범위의 출현을 보였다. 이매패류인 애기반투명조개 Theora lata는 비교적 남해 해역에 위치한 정점들에서 우점 출현하였고, 전체적으로 연안의 육지에 인접한 정점들에서 높은 밀도로 나타났다.
Table 3.
Major dominant species of macrobenthos in Korean coastal waters in August 2017
| Species name |
Mean Density (ind./m2) | % | Frequency |
| Pseudopolydora kempi (P) | 313 | 25.5 | 8 |
| Heteromastus filiformis (P) | 84 | 6.8 | 87 |
| Magelona johnsoni (P) | 54 | 4.4 | 25 |
| Lumbrineris longifolia (P) | 38 | 3.1 | 60 |
| Dorvillea matsushimaensis (P) | 31 | 2.5 | 17 |
| Theora fragilis (B) | 29 | 2.4 | 46 |
| Sternaspis scutata (P) | 27 | 2.2 | 66 |
| Praxillella affinis (P) | 25 | 2.0 | 40 |
| Maldane cristata (P) | 20 | 1.7 | 16 |
| Sinocorophium sinensis | 19 | 1.6 | 18 |
| Rhynchospio sp. (P) | 19 | 1.5 | 1 |
| Tharyx sp. (P) | 17 | 1.4 | 74 |
| Ampelisca misakiensis (C) | 13 | 1.1 | 30 |
| Eriopisella sechellensis (C) | 13 | 1.0 | 34 |
| Aricidea jeffreysii (P) | 12 | 1.0 | 50 |
3.4 집괴분석과 정점군 특성
한국 연안해역의 117개 정점에서 채집된 대형저서동물의 모든 개체수 자료를 바탕으로 집괴분석을 수행하였다. Bray-Curtis 유사도 20%를 기준으로 정점을 분류한 결과 지리적으로 구분되는 6개의 정점군이 분류되었다(Table 4, Fig. 4). 정점군 A는 수심이 깊고 사질 함량이 낮은 정점 10개로 구성되며 동해 해역에 위치한다. 출현종수는 정점군 B와 C에 비해 적지만 평균 서식밀도는 가장 많은 2,033 개체/m2였다. 우점종은 양손갯정이류인 M. johnsoni와 송곳갯지렁이류인 L. longifolia였으며, 이 두 종이 전체 개체수의 42.6%를 차지하였다. 정점군 B는 수심이 약 20 m 정도이고 실트 함량이 50% 이상이며, 유기물 함량이 높은 정점들로 구성되며 남해 해역에 위치한다. 출현종수는 다른 정점군에 비해 가장 많은 407종이며 평균 서식밀도는 두 번째 높은 밀도인 1,279 개체/m2였다. 정점군 C는 다른 정점군에 비해 수심이 가장 낮은 약 20 m 이하이고 사질 함량이 약 50% 이상인 정점들로 구성되며 서해 해역에 위치한다. 출현종수는 정점군 B 다음으로 높아 287종이 출현하며, 우점종은 버들갯지렁이류인 H. filiformis로서 전체 개체수의 28.2%를 차지하였다. 정점군 D는 정점군 B와 유사한 수심인 약 20 m 정도이고 실트 함량이 약 60% 이상이었으며, 유기물 함량이 다른 정점군에 비해 가장 높은 값을 보였다. 정점군 D는 서해남부 해역에 위치하며, 우점종인 애기반투명조개 T. lata는 전체 개체수의 40.4%로 매우 높은 비율을 차지하였다. 정점군 E는 수심이 약 23 m 정도이고 사질 함량이 약 95% 정도이며 서해중부 해역에 위치한다. 출현종수는 86종으로 정점군 D와 F를 제외하고 가장 적었으며 서식밀도는 다른 정점군에 비해 가장 낮은 값을 보였다. 정점군 F는 수심이 약 30 m 정도이고 사질함량이 약 97% 정도로 높은 값을 보였다. 유기물 함량은 다른 정점군에 비해 가장 낮은 값을 보였으며, 서해북부 해역에 위치한 정점들로 이루어졌다.
Table 4.
Characteristics of benthic environment and macrobenthic community of each group divided by cluster analysis in Korean coastal waters, August 2017
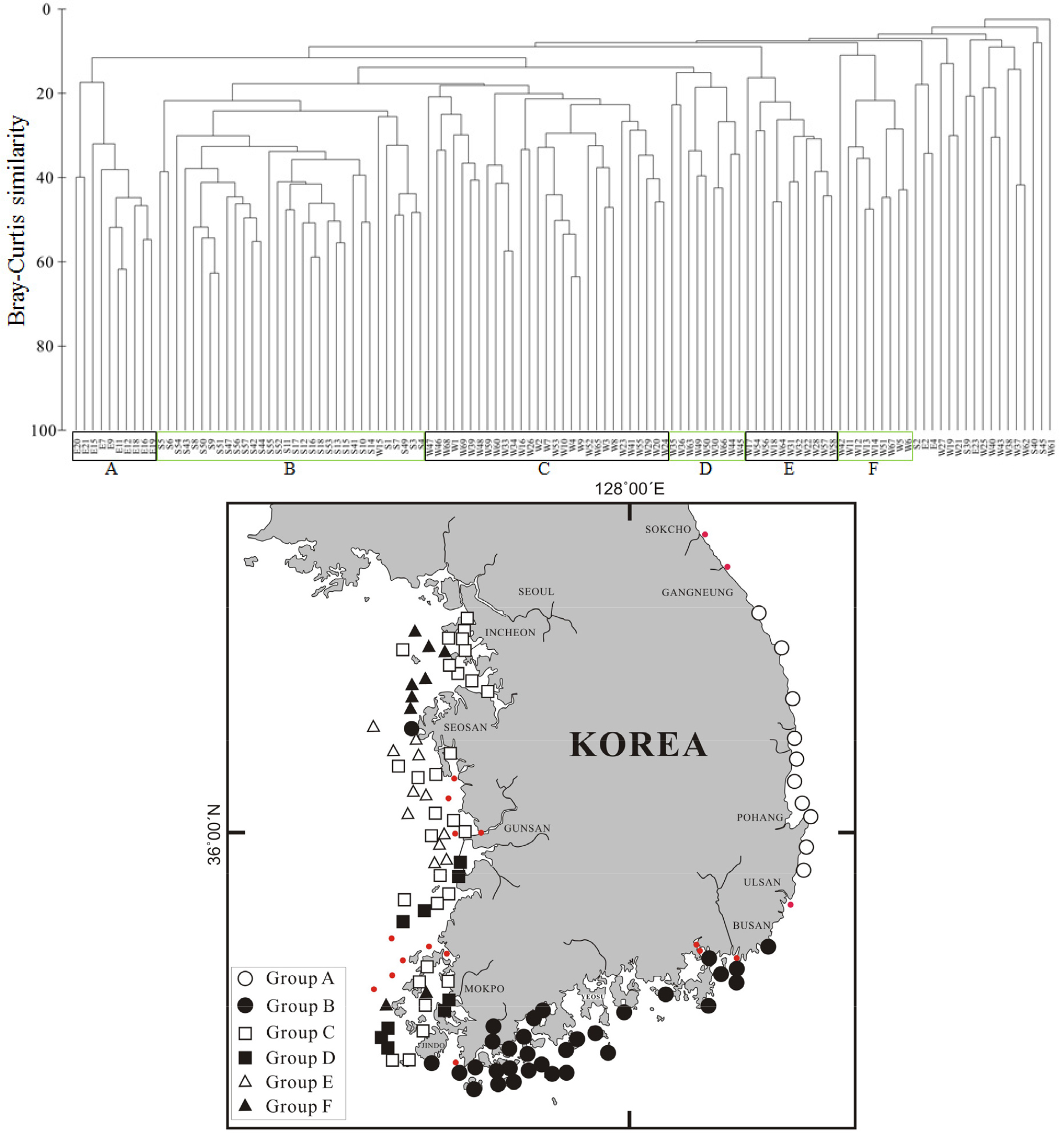
집괴분석을 통해 6개 정점군의 유사도에 영향을 주는 생물을 파악하기 위해 SIMPER 분석을 수행하였다(Table 5). 정점군 A는 전체 유사도 26.9% 이였으며, 정점군의 상위 우점종과 동일한 종들이 기여도에 영향을 주었다. 정점군 B는 상위 우점종과 다른 단각류인 E. sechellensis (9.39%)와 오뚜기갯지렁이인 S. scutata (8.69%)가 기여도에 영향을 주었다. 정점군 C,D,E,F는 최상위 우점종이 다른 우점종에 비해 높은 기여도를 차지하였다.
Table 5.
SIMPER analysis of macrobenthos listing the main contribution species at each groups
또한 대형저서동물의 군집구조에 영향을 미치는 주요 환경요소을 파악하기 위해 Bio-Env 분석 결과 표층퇴적물의 평균입도, 분급도, 수심과 유기물함량 요소가 복합적으로 영향을 미치는 것으로 나타났다(Bio-Env: r=404).
3.5 상관관계 및 주성분분석
한국 연안에 분포하는 대형저서동물 군집 요소와 저서환경 요인간의 상관관계를 알아보기 위해 Spearman rank correlation coefficent를 분석한 결과는 Table 6과 같다. 출현종수는 표층퇴적물의 분급도, Clay, Silt, 유기물 함량(LOI)과 높은 양의 상관관계를 보였으며, Sand와는 음의 상관관계를 보였다. 생물량과 풍부도지수는 출현종수와 비슷한 양상의 상관관계를 보였다. 우점종인 양손갯지렁이류 M. johnstoni, 긴자락송곳갯지렁이 L. longifolia와 대나무갯지렁이류 M. cristata는 수심, 총유기탄소량(TOC)과 높은 양의 상관관계를 보였다. 애기반투명조개 T. lata와 옆새우류 E. sechellensis는 평균 입도, Silt, Clay, 유기물 함량(LOI), 총유기탄소량(TOC)과 높은 양의 상관관계 보였으며, Sand와는 음의 상관관계를 보였다.
우점종과 저서환경 요인간의 상관관계를 보면 Sand 함량과 우점종인 T. lata와 E. sechellensis는 비교적 높은 음의 상관관계를 보였으나 다른 우점종들은 낮은 상관관계를 보였다. 유기물 함량과 우점종인 T. lata, S. sinensis와 E. sechellensis은 비교적 높은 양의 상관관계를 보였으나 다른 우점종들은 낮은 상관관계를 보였다.
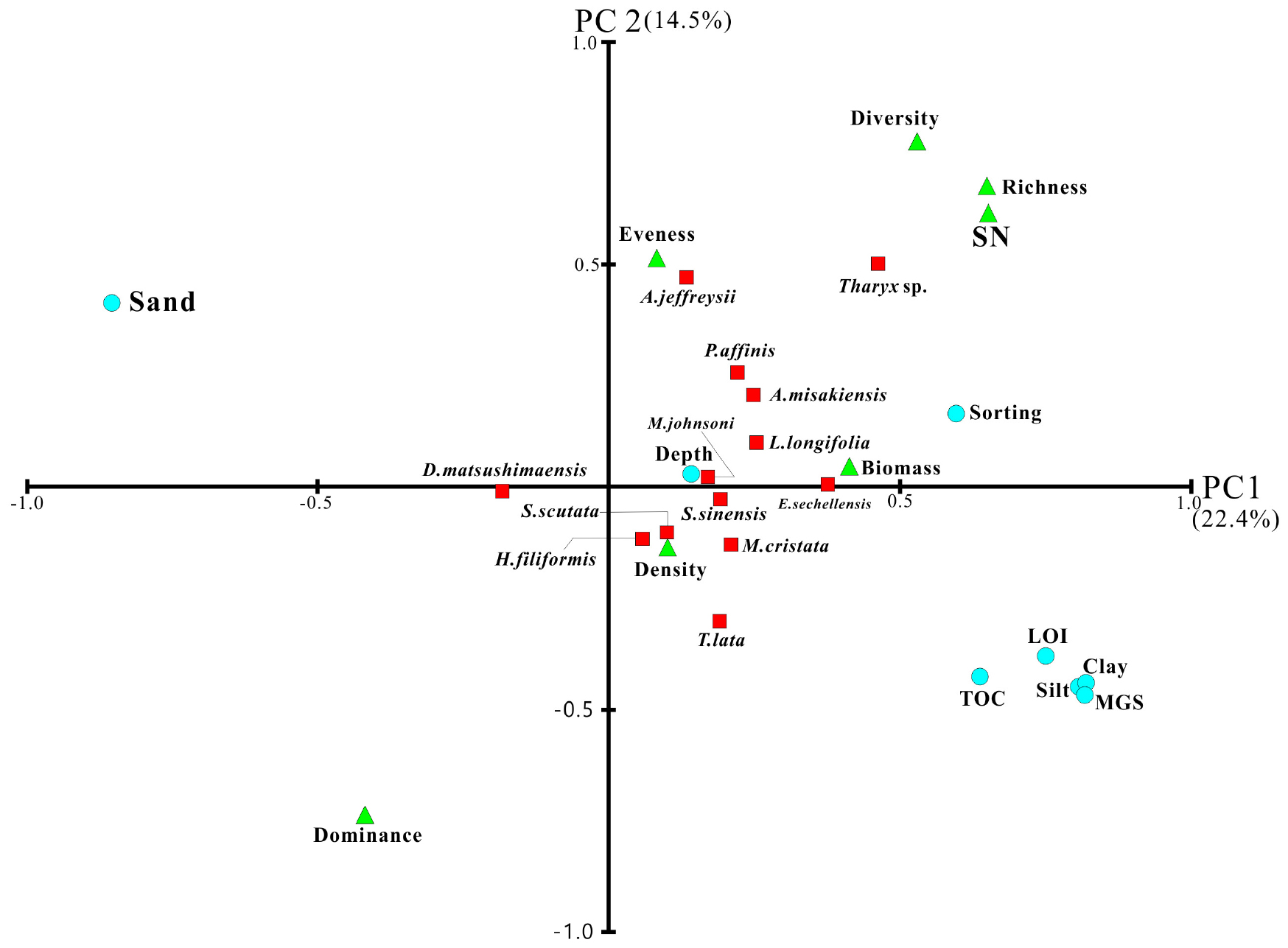
한국 연안의 저층환경 요인과 대형저서동물 군집를 파악하기 위해 저층환경 요인 8개와 우점종 13종, 생태지수를 대상으로 주성분분석을 하였다(Fig. 5). 주성분 1은 전체 분산의 22.4%의 기여율을 보였으며, 평균입도, Clay, Silt, LOI는 상관계수 0.7 이상과 Sand는 상관계수 –0.8 이상의 상관관계를 보이고 있어 저서퇴적환경을 의미한다고 해석할 수 있다. 주성분 1은 저서퇴적환경과 출현종수(r=0.651), 풍부도지수(r=0.648), 다양도지수(0.529)간에 높은 상관관계를 보이고 있으며, 이는 니질함량이 증가할수록 출현종수는 많아지는 양상을 보인다. 우점종인 실타래갯지렁이류 Tharyx sp. (r=0.462)와 옆새우류 E. sechellensis (r=0.378)의 서식밀도는 저층퇴적환경과 비교적 높은 양의 상관관계를 보였다. 주성분 2는 전체 분산의 14.5%의 기여율을 보였고 환경요인과 상관관계를 나타내지 않았다. 우점도(r=-0.742)는 음의 상관관계를 보였고 다양도(r=0.765), 풍부도(r=0.666), 출현종수(r=0.610)는 양의 상관관계를 보였다. 이러한 결과로 보아 대형저서동물의 출현종수가 많아지면 우점도 지수는 낮아지는 것을 의미한다.
Table 6.
Spearman rank correlation coefficients between benthic environmental factors and dominant species in Korean coastal waters
| S.N | Density | Biomass | R | E | D | Do | H. filiformis | M.johnsoni | L.longifolia | D.matsushimaensis | T.lata | S.scutata | P.affinis | M.cristata | S.sinensis | Tharyx sp. | A. misakiensis | E.sechellensis | A.jeffreysii | |
| MGS | .264** | .236* | .332** | .247** | -0.075 | 0.181 | -0.102 | .229* | 0.102 | .371** | -0.023 | .479** | .393** | .206* | .207* | .357** | 0.148 | .226* | .422** | -0.135 |
| Sorting | .386** | .313** | 0.173 | .387** | 0.059 | .303** | -.255** | .427** | -0.139 | .341** | 0.038 | .251** | .204* | .412** | 0.094 | 0.093 | .301** | 0.161 | 0.162 | .336** |
| Sand | -.290** | -.251** | -.303** | -.276** | 0.054 | -.197* | 0.106 | -.261** | -0.045 | -.369** | -0.023 | -.440** | -.308** | -.261** | -0.172 | -.365** | -0.142 | -.287** | -.384** | 0.110 |
| Silt | .258** | .224* | .267** | .245** | -0.061 | 0.173 | -0.098 | .256** | 0.062 | .356** | -0.021 | .424** | .373** | .247** | 0.178 | .401** | 0.127 | .269** | .396** | -0.104 |
| Clay | .304** | .274** | .348** | .288** | -0.077 | .201* | -0.118 | .269** | 0.059 | .372** | -0.011 | .469** | .324** | .240** | .210* | .311** | 0.171 | .239** | .393** | -0.080 |
| Depth | 0.178 | 0.128 | .210* | 0.175 | 0.032 | .192* | -0.172 | -0.172 | .285** | 0.170 | .374** | -.194* | -0.176 | .225* | .432** | -0.137 | 0.144 | 0.052 | -0.169 | -0.077 |
| LOI | .322** | .247** | .326** | .316** | -0.043 | .239* | -0.138 | .273** | 0.016 | .414** | 0.057 | .469** | .362** | .280* | .203* | .353** | .218* | .225* | .441** | -0.026 |
| TOC | .307** | .286** | .319** | .290** | -0.084 | .195* | -0.095 | 0.173 | 0.182 | .457** | .199* | .388** | .285** | .322** | .372** | .310** | .235* | 0.140 | .295** | -0.068 |
4. 고 찰
4.1 대형저서동물 종 특성
한국의 전 연안해역을 대상으로 대형저서동물을 조사한 결과는 전무한 실정이며, 본 조사는 여름철에 117개 정점에서 조사된 결과 총 613 종/35.1 m2, 평균 1,228 개체/m2의 대형저서동물이 채집되었다. 우리나라 연안에서 그동안 수행된 대형저서동물 군집 연구 결과들 가운데 남해도 외해역에서 계절별로 조사된 247종, 1,026 개체/m2 (Lim et al., 2016), 3년동안 낙동강 하구역에서 조사한 266종, 859 개체/m2 (Seo et al., 2016), 서해 해역인 천수만에서 계절별로 조사한 311종, 769 개체/m2 (Park et al., 2000) 등과 비교해 보면 본 조사 결과는 출현종수 및 서식밀도가 높은 것으로 나타났다. 그리고 여름철 동해 해역에서 조사된 253종, 4,895 개체/m2 (Kwon et al., 2017), 여름철 남해 해역인 여자만에서 조사된 274종, 2,346 개체/m2 (Lim, 2015)과 비교해 보면, 출현종수는 많았으며 서식밀도는 낮게 나타났다(Table 7).
Table 7.
Comparison of macrobenthic sommunity structures studied in castal regions of Korea
| Study area | Depth (m) | Number of species | Density (ind./m2) | Sampling Season | Sampling area (m2) | Refernces |
| Nambaedo Is. (S) | 10-30 | 247 | 1,026 | seasonal | 16.0 | Lim et al.(2016) |
| Nakdong estuary (S) | 10-40 | 266 | 859 | 3 yrs. | 19.2 | Seo et al.(2016) |
| Chonsu Bay (W) | <20 | 311 | 769 | seasonal | 25.2 | Park et al.(2000) |
| Uljin area (E) | 10-65 | 253 | 4,895 | summer | 4.6 | Kwon et al.(2017) |
| Yoja Bay (S) | <20 | 274 | 2,346 | summer | 17.4 | Lim(2015) |
| Castal regions of Korea | <90 | 613 | 1,228 | summer | 35.1 | This presemt study |
본 연구 결과와 채집 시기 및 횟수, 정점수, 기간 등의 차이로 인해 다른 조사 결과와 직접적인 비교는 어렵지만, 출현종수는 채집면적, 기간, 정점수등이 증가할수록 희소종의 추가로 인해 증가할 수 있다. 따라서 다른 조사해역 보다 채집면적의 증가로 인하여 출현종수가 증가한 것으로 해석된다. 그리고 여름철 1회성 조사임에도 불구하고 출현종수가 많은 것은 동해는 수심이 깊고 서해는 담수유입이 많고 남해는 니질함량 높은 지역으로 다양한 서식환경으로 인하여 출현종수 증가에 영향을 준 것으로 판단된다. 이러한 결과는 다양한 서식지 환경조건(입도, 염분, 수온, 수심 등)을 포함할수록 다양한 저서동물이 출현하게 된다(Lim, 2015)는 결과와도 일치한다.
대형저서동물의 서식밀도 증가는 상위 우점종의 총 출현 개체수 비율에 의해 결정되는 것으로 해석해 볼 수 있다. 서식밀도가 높은 여자만(2,346 개체/m2)과 동해해역(4,895 개체/m2)은 최우점종의 개체수 비율은 각각 49.7%, 71.0%를 차지하여 한 종이 높은 밀도로 극 우점하였으며, 본 조사(1,228 개체/m2) 및 남해도(1,026 개체/m2)해역에서는 각각 8종, 7종이상 채집되어야만 개체수 비율이 50%이상을 차지하는 것으로 나타났다. 이러한 결과는 상대적이긴 하나, 총 출현 개체수 비율이 한,두종의 극 우점 보다는 다양한 종들의 균등한 출현이 밀도 감소에 영향을 주는 것으로 판단된다. 또한 국한된 조사해역의 특징종이 출현함으로써 그 해역의 밀도 증가에 영향을 주는 것으로 해석해 볼 수 있다. 여자만은 기수역에 출현하는 종(Minuspio japonica; 1,167 개체/m2)과 동해해역은 사질퇴적상에서 출현하는 종(Spiophanes bombyx; 3,031 개체/m2)으로 인하여 전체적인 서식밀도에 영향을 주는 것으로 해석된다. 따라서 우리나라 연안해역의 밀도 조성은 다양한 종들이 우점 출현하고 특징종의 극 우점한 경향이 나타나지 않은 결과로 판단된다.
4.2 주요 우점종 특성
본 조사에서 채집된 대형저서동물의 상위 우점종은 대부분 환형동물이 차지하였으며, 첫 번째 우점종인 선녀얼굴갯지렁이류 Pseudopolydora kempi는 낙동강 하구 입구 해역인 정점 45번에서만 약 36,000 개체/m2로 극우점 출현하는 특징을 보였다. 이 종은 국외 및 국내에서도 유기물 오염이 진행된 해역에서 우점하는 종으로 알려져 있는데 내만 해역인 시화호, 인천북항, 영일만과 낙동강 등에서 우점 출현하는 것으로 보고되었다(Pearson and Rogenberg, 1978; Shin et al., 1992; Ryu et al., 1997; Ryu, 1997; Seo et al., 2016). Seo et al.(2016)에 의하면 낙동강 하굿둑 방류량에 따라서 P. kempi가 대량 유입된다고 보고하고 있어 본 조사 시기가 여름철인 점을 감안하면 담수가 대량 유입되어 본 종은 퇴적물과 함께 유입 되었을 것으로 판단된다.
두 번째 우점종인 고리버들갯지렁이 Heteromastus filiformis도 유기물 오염이 심한 해역에서 높은 밀도로 출현하는 지표종으로 많이 보고된 종이다(Grizzle, 1984; Tsutsumi et al., 1991; Lim et al., 2006; Jang and Shin, 2016). 본 조사에서는 서해와 남해 해역의 육지에 인접한 해역이나 하구역에서 우점 출현하였으나 H. filiformis 서식 밀도와 퇴적물 유기물 함량(LOI) 및 총유기탄소량(TOC) 사이에는 상관관계가 크지 않았다(LOI; r=0.043, TOC; r=-0.009). 내만 해역이나 육지에 인접한 해역에 국한된 조사에서는 높은 유기물 함량에서 H. filiformis가 우점 출현하고 있다. 그러나 우리나라 연안의 다양한 환경(수심, 염분, 수온, 용존산소 등)에서는 유기물 오염이 되지 않은 해역에서도 밀도가 높게 나타나거나 유기물 오염 해역에서 밀도가 적게 나타나는 경우도 있어 이 종의 생태학적 특성에 대해 보다 상세한 연구가 필요하다고 판단된다.
세 번째 우점종인 양손갯지렁이류 Magelona johnstoni는 동해 해역에서만 높은 밀도로 출현하는 양상을 보였으며, 수심(r=0.321) 및 총유기탄소량(r=0.290)과 비교적 높은 양의 상관관계가 나타났다. 이러한 결과로 보아 이 종은 수심이 깊고 퇴적물의 유기 탄소가 많은 해역에서 높은 서식밀도를 보이는 것으로 판단된다. 그리고 동해연안 해역의 이전 연구에서는 속명이 동일한 Magelona sp.가 1985~987년(Choi and Koh, 1990), 2005년(Paik et al., 2007), 2012~2013년(Kwon et al., 2017) 및 2013~2016년(Bae et al., 2018)에 우점 출현하는 것으로 보고되었다. 이전 연구의 Magelona sp.는 표본이 보관되어 있지 않아서 정확한 형태적 특징을 확인하기 어려우나 동일한 종으로 추정된다. 향후 분류학적으로 동일종임이 확인된다면 동해 해역의 대형저서동물군집 생태계 변동 연구에 중요한 생물종으로 판단된다.
긴자락송곳갯지렁이 Lumbrineris longifolia는 H. filiformis와 마찬가지로 유기물 오염이 된 해역에서 높은 밀도로 출현하는 것으로 보고되었다(Lim et al., 1992; Shin, 1995; Baik and Yun, 2000). 본 조사에서도 총유기탄소량과 L. longifolia 서식밀도는 양의 상관관계가 유의한 수준으로 나타났다(r=0.316). 또한 수심이 깊을수록 밀도가 높게 나타나는 상관관계를 보였으며(r=0.260), 이는 반폐쇄적인 내만이나 육지에 인접한 해역뿐만 아니라 수심이 깊은 해역에서도 퇴적물의 유기물량이 많다면 높은 밀도로 출현할 수 있음을 시사한다.
4.3 군집구조 및 상관분석
본 조사에서 출현한 대형저서동물의 종별 개체수 자료를 이용하여 집괴분석한 결과 크게 6개 정점군으로 구분되었다. 즉, 우리나라 연안의 대형저서동물군집은 지리적으로 3개의 해역 동해 해역(A 정점군), 남해 해역(B 정점군), 서해 해역(C 정점군)으로 구분되었고 나머지 정점군들은 서해 해역이 북부 외해, 중부, 남부 해역으로 구분되었다(Table 4). 대형저서동물군집은 다양한 저서환경 변화(유기물 농도, 염분 구배, 용존산소, 입도, 수심)로 인하여 영향을 받아 군집이 구분되어진다고 보고되었다(Gray, 1974; Park et al., 2000; Yoon, 2009; KORDI, 2012; Kim et al., 2016; Seo et al., 2016; Lee and Ryu, 2018). 따라서 본 연구 해역의 군집분석 결과도 기존 결과들과 유사한 양상을 나타낸다. 동해 해역은 남해 해역과 비슷한 입도조성과 유기물 함량을 보이고 있으나 수심에서 큰 차이를 보였으며, 남해 해역은 서해 해역과 비슷한 수심을 보이고 있으나 사질 함량에서 차이를 보였다. 이러한 결과로 보아 동해 해역은 깊은 수심과 많은 유기물 함량, 적은 사질 함량이 군집에 영향을 미치는 주요인으로 판단된다. 그리고 남해 해역은 낮은 수심과 많은 유기물 함량, 적은 사질 함량이 영향을 미치고 있으며, 서해 해역은 낮은 수심과 많은 사질 함량 및 유기물 함량으로 인하여 종조성의 변화가 나타났고 이로 인하여 군집의 차이가 나타났다고 판단된다(Table 4).
우점종들과 저서환경 요소들 사이에는 상관관계가 뚜렷하게 나타나고 있으나(Table 4, 5), 우리나라 연안에서 니질 함량이 높고, 유기물 함량이 높은 해역에서 흔히 볼 수 있는 버들갯지렁이류 H. filiformis 및 오뚜기갯지렁이 Sternaspis scutata는 저서환경요인과 뚜렷한 상관관계를 보이지 않았다. 이는 우점종 분포에서 언급한 바와 같이 두 종에 대한 생태학적 특징을 다양한 관점에서 검토할 필요성이 제기된다.
결론적으로 우리나라 연안의 대형저서동물군집의 공간 분포는 수심, 퇴적상, 유기물 함량에 의해 종조성이 변화하고 있으며, 이로 인하여 정점군이 구분되어지는 것을 알 수 있었다. 이것은 우리나라 내만역이나 반폐쇄적인 내만에서 연구된 대형저서동물군집 분포와 비슷한 결과로서 결국, 우리나라 연안의 대형저서동물 군집 분포에 영향을 주는 저서환경 요소들은 지금까지 언급된 범주를 크게 벗어나지 않는다고 할 수 있다.
