

1. 서 론
2. 자료 및 방법
2.1 자료
2.2 자료분석 방법
2.3 평가 프로그램
3. 결 과
3.1 어획물 생물량과 종조성
3.2 제주도 남서부해역 어획 어종 공간분포 특성
3.3 그룹별 생물량 비교
3.4 그룹별 생태학지수
3.5 그룹별 우점종 체장조성
4. 고 찰
1. 서 론
동중국해의 북동부에 위치하는 제주도 근해 해역은 동중국해와 우리나라 남해, 황해를 연결하는 해역으로 쿠로시오난류에서 분기된 대마난류수, 황해 저층냉수와 여름철 중국 양자강 유역에서 유입되는 중국대륙 연안수 등 성질이 다른 수괴의 영향으로 계절별 복잡한 수문학적 특성을 나타낸다(Moon et al., 2009). 이 중 제주도 남서부 연근해 해역은 황해 남부와 동중국해 북부 일부가 포함되며, 수심 200 m 미만의 넓은 대륙붕과 좁은 대륙사면으로 이루어진 해역이다(Lee et al., 2022). 또한, 연중 10℃ 이상의 난류수의 형성과 풍부한 먹이생물 등의 조건으로 각종 회유성 어류들의 주요 산란장이자 월동장이기도 하며, 우리나라 주변 해역 중 어선어업이 활발한 좋은 어장으로 어획 생산량이 높아 경제적으로 중요한 해역으로 구분될 뿐만 아니라 기후변화에 가장 민감하게 반응하는 해역으로 구분된다(Cha et al., 2001; Heo et al., 2015; Kim, 2017).
우리나라 주변해역 연근해 종조성 평가 연구는 일반적으로 중・저층 트롤과 같은 대형어구로 수행되었다(Cha and Park, 1747; Huh and Kwak, 1748; Koh et al., 2016). 이 중 저층 트롤은 조업의 대상이 되는 어종과 비대상 해양생물을 함께 어획하여 혼획의 위험이 높은 어구로 구분되지만 다른 어구와 비교해서 어획성능이 뛰어나고 이동성이 적은 불특정 다수 생물을 선택성 없이 어획할 수 있는 장점 때문에 특정해역의 생물상을 조사하는 연구에 주로 사용되고 있다(Lee, 2011; Lee et al., 2015).
최근까지 진행된 제주도 주변해역 연근해 수산자원 종조성 평가에 대한 연구로 Kim and Kim (2010)의 한일공동수역의 저서 어획물 분포특성 연구와 Shin et al. (2023)의 제주도 남서부 해역의 수산자원 종조성과 밀도를 평가하기 위한 연구가 수행되었다. 저층 트롤을 활용한 우리나라 주변 해역 해양생물 종조성 평가 연구는 대부분 연안과 만을 중심으로 수행되었다(Han and Oh, 2007; Kim et al., 2007a; Choi et al., 2014; Koh et al., 2015; Joo et al., 2020). 저층 트롤을 활용한 근해 조사는 황해 중앙부와 동해 심해조사(수심 250~1,030 m), 부산-대마도 경계수역, 제주도와 남해 사이의 근해와 제주도 동부해역에 한정하여 수행되어 우리나라 주변 해역 중 어업 생산량이 가장 높고 해양환경 변동에 가장 우선적으로 반응하는 제주도 남서부 해역에 대한 해양생물 종조성과 분포 연구는 상대적으로 부족한 실정이다(Jeong et al., 2005; Park et al., 2007; Kim et al., 2010; Kim et al., 2011; Koh et al., 2016; Kim et al., 2018; Joo et al., 2020). 또한, 외해의 해양생물 종조성 평가를 위한 조사는 조사선 확보와 트롤 장비 같은 대형 어구를 운영하기 위한 높은 경제적 비용이 요구되기 때문에 주로 국가 연구기관이나 대학을 중심으로 수행되고 있으며, 어선이나 정치성 어구를 활용한 연안 조사보다 자료 확보가 어려워 외해역 해양생물 종조성에 대한 이해는 아직 미흡한 수준에 머물러 있다.
따라서, 본 연구는 제주도 남서부 외해를 포함한 연근해 해역을 대상으로 저층 트롤을 이용한 어획 시험 조사 수행하여 여름 해양생물의 종 다양성과 공간 분포 특성을 파악하고 조사 정점별(Station, St) 생물종 구성과 생물량을 기준으로 정리하여 공간별 분포하는 해양생물의 차이와 정점별(St) 특성을 비교하고자 한다. 본 연구 결과는 추후 수행될 한국 남해 어획 시험조사와 연결하여 여름 북동중국해 해양생물 종조성을 확장하여 해석하기 위한 자료가 될 것이다.
2. 자료 및 방법
2.1 자료
본 조사는 2020년 7월과 8월 총 4회에 걸쳐 전남대학교 새동백호를 이용하여 이루어졌으며, 트롤 조사에 사용된 트롤 어구의 규격은 Fig. 1와 같다. 조사에 이용된 저층 트롤 그물의 총 길이는 50.2 m이었으며, 뜸줄의 길이는 35.2 m, 발줄의 길이는 45.6 m, 그물의 높이와 폭은 2.5 m와 26.5 m이다. 끝자루 길이는 8.6 m이며, 끝자루의 높이는 1.5 m, 끝자루의 폭은 2.2 m이다. 끝자루 그물은 결절 그물감을 사용했으며, 그물의 안쪽 그물코 크기는 120 mm, 바깥쪽 그물코 크기는 240 mm로 구성되었다(Shin et al., 2023).
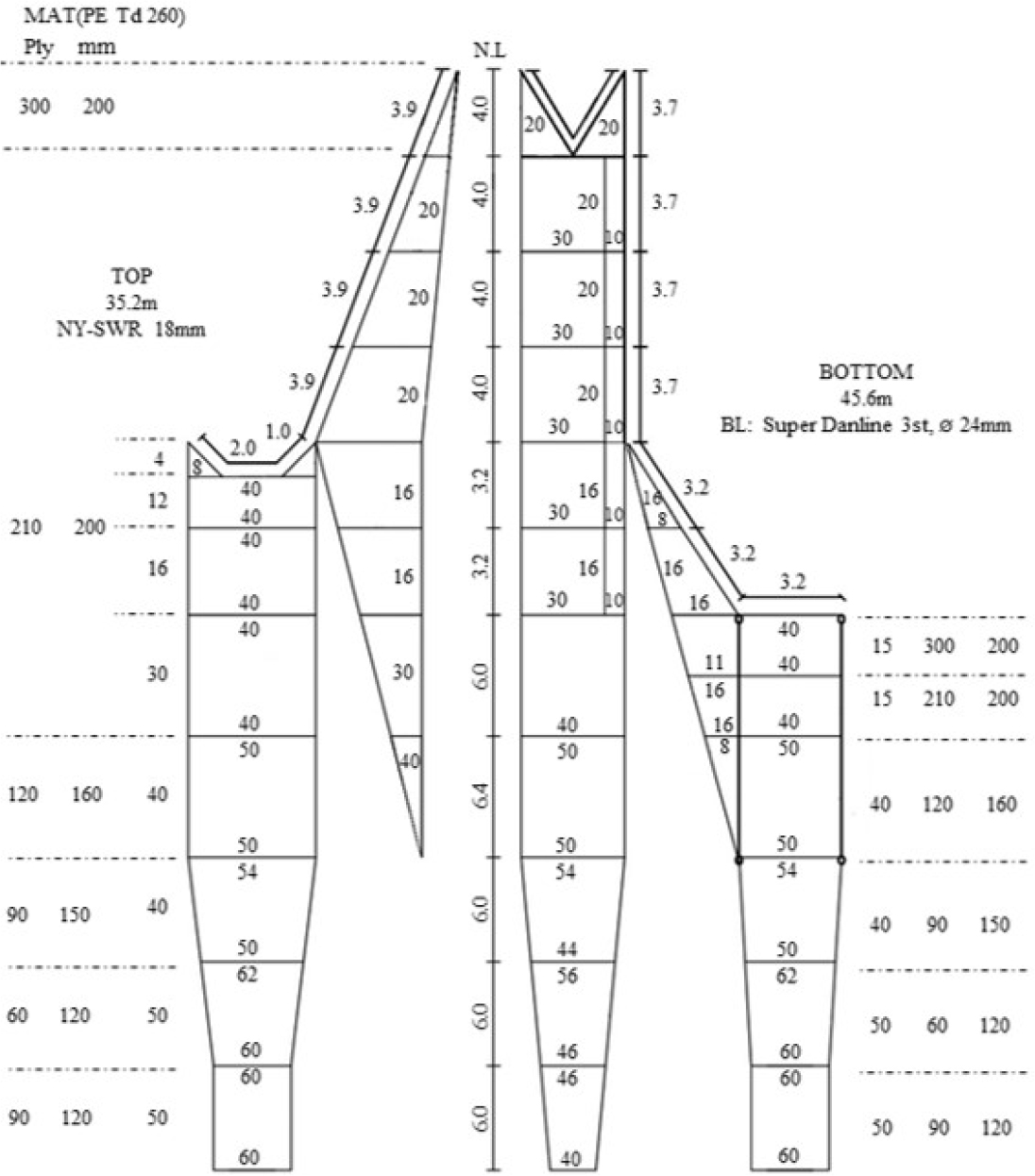
조사해역은 Fig. 2와 Table 1에 나타난 총 5개 지점에서 이루어졌으며, 정점별(St) 채집 수심은 75.5~107.5 m, 예망 속도는 3.2~3.6 knot로 수행하였고, 평균 예망시간은 약 30분 으로 수행하였다. 어획물은 양망 후 어종별로 분류하여 어종별 개체수를 파악하였으며, 어종이 구분된 개체는 전장을 기준으로 1 cm, 중량은 저울을 이용하여 1 g단위로 측정하였다. 개체의 분류는 형태적으로 분류한 이후 Froese and Pauly(2020)와 Yamada et al.(2007)를 기준으로 과수준의 분류를 수행하였다.
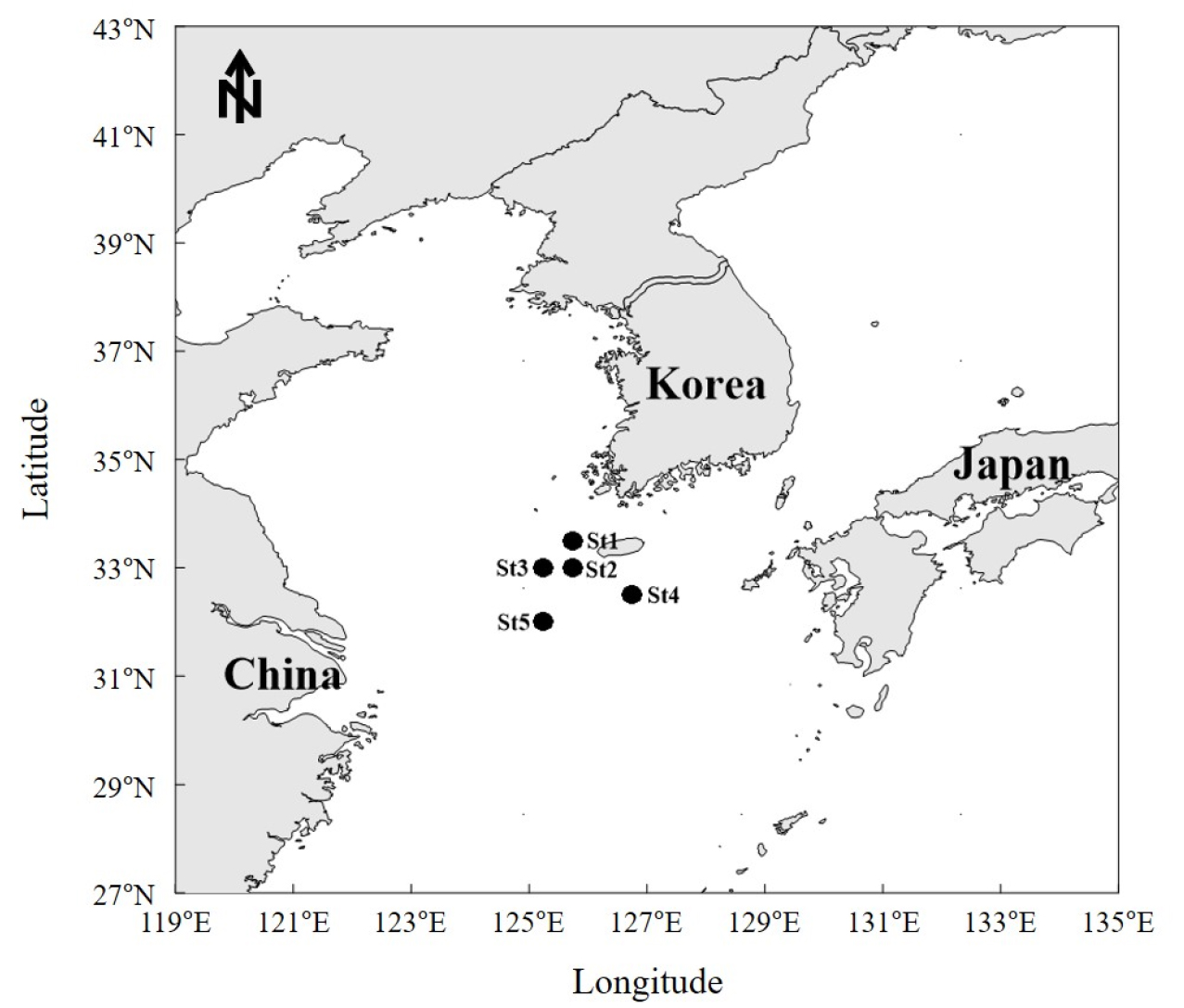
Table 1.
The information on the survey area including latitude, longitude, date and depth for each station
2.2 자료분석 방법
어종별 공간 분포는 어획량 상대 비율 변화를 고려하여 군집의 분포를 평가하는 대응분석(CA, Correspondence analysis)으로 평가하여 그룹별로 요약하였으며(Ludwig and Reynolds, 1988), 총 어획량의 1%를 기준으로 우점종을 구분하고 생물량과 체장조성을 확인하였다.
그룹별 어획어종 군집 특성을 파악하기 위해 종 다양성은 Shannon and Weaver(1963)의 종 다양도지수(Species diversity index, H’), Margalef(1969)의 종 풍부도지수(R), Pielou(1966)의 균등도지수(J)와 McNaughton(1968)의 우점도지수(D)를 계산하였으며, 그 식은 아래와 같다.
여기서, pi는 i번째 속하는 어종의 개체수 비율(ni/N)을 의미하며, S는 출현 종 수, N은 총 출현 개체수, Y1과 Y2는 첫 번째와 두 번째 우점종 개체수를 의미한다. 그룹별 우점종의 성숙 개체 분포 특성은 어종별 체장빈도를 과거 문헌으로 평가한 군성숙체장(L50, 50% group maturity length)과 비교하여 평가하였다.
2.3 평가 프로그램
종 다양도지수(H’), 종 풍부도지수(R), 균등도지수(J)와 우점도지수(D)는 액셀 2019 프로그램을 사용하여 계산하였으며, 공간분포 평가를 위한 대응분석은 Ludwig and Reynolds (1988)가 제시한 방법에 따라 R 통계프로그램(4.0.4)으로 수행하였다.
3. 결 과
3.1 어획물 생물량과 종조성
2020년 7월과 8월 총 4회의 저층 트롤 조사를 수행하여 어획된 해양생물은 어류 23과 27종, 688개체 158,453 g이 어획 되었으며, 갑각류는 6과 6종 274개체, 13,016 g, 두족류는 3과 3종 63개체 8,920 g과 복족류 1과 1종 1개체 150 g가 어획되었다(Table 2). 이 중 황아귀(Lophius litulon)가 총 어획 생물량의 53.04%를 차지하여 가장 많은 것으로 나타났으며, 다음으로 민달고기(Zenopsis nebulosa) 8.28%, 꼼치(Liparis tanakae) 4.15%, 전갱이(Trachurus japonicus) 3.76%, 붉은메기(Hoplobrotula armata) 3.33%, 넙치(Paralichthys olivaceus) 3.32%, 홍어(Okamejei kenojei) 2.66%, 달고기(Zeus faber) 2.63%, 살오징어(Todarodes pacificus) 2.54%, 두드럭게(Leptomithrax edwardsii) 2.18%, 부채새우(Metanephrops thomsoni) 2.17%, 창꼴뚜기(Loligo edulis) 1.91%, 가시발새우(Solenocera melantho) 1.69%, 눈볼대(Doederleinia berycoides) 1.19%가 차지하였으며, 나머지 종 들은 총 어획개체의 1% 미만으로 나타났다(Table 2).
3.2 제주도 남서부해역 어획 어종 공간분포 특성
종별 어획 생물량을 기준으로 공간분포 특성을 평가하기 위한 대응분석을 수행한 결과 세 그룹으로 구분되었다(Fig. 3). 어획 생물량 기준 그룹별 우점종을 구분하면, 그룹 1 (St1, St3, St5)에서는 황아귀, 꼼치, 넙치가 우점하는 것으로 나타났으며, 그룹 2 (St2)는 달고기, 전갱이, 황아귀, 대롱수염새우(Solenocera melantho), 홍어, 붉은메기, 그룹 3 (St4)은 황아귀, 달고기, 붉은메기, 두드럭게, 창꼴뚜기, 살오징어 순으로 생물량이 높게 나타났다(Table 3).

Fig. 3.
Correspondence analysis of biomass composition of the dominant 11 species from survey data in the southwestern sea of Jeju Island in Summer with respect to fish species (a) and stations (b). The first (42.1%) and second (28.7%) dimensions explained the variability in biomass composition by species. (a) and (b) can be overlapped for graphical interpretation.
3.3 그룹별 생물량 비교
대응분석을 통해 구분한 그룹별 어획물의 총 개체수와 생물량은 Fig. 4와 같다. 그룹별 총 개체수(n)는 그룹 1 에서 573개체로 가장 높게 나타났으며, 그룹 2 259개체, 그룹 3은 195개체 순으로 나타났다. 그룹별 어획 생물의 총 생물량(g)은 그룹 1 에서 114×103의 가장 높은 생물량을 나타냈으며, 그룹 2는 39×103, 그룹 3은 29×103 순으로 나타났다. 그룹별 어획 생물은 개체수와 생물량 모두 그룹 1에서 가장 높으며, 다음으로 그룹 2, 그룹 3 순으로 나타났다.

3.4 그룹별 생태학지수
대응분석을 기반으로 제주도 남서부해역의 그룹별 생태학지수는 Fig. 5와 같다. 종 다양성지수(H’)는 1.91~2.28의 범위로 그룹 3이 가장 높고, 그룹 1이 가장 낮게 나타났다(Fig. 5a). 종균등도지수(R)는 3.22~6.3의 범위로 그룹 1이 가장 높고, 그룹 3에서 가장 낮게 나타났다(Fig. 5b). 종우점도지수(J)는 0.51~0.79의 범위로 그룹 3에서 가장 높고 그룹 1에서 가장 낮게 나타났다(Fig. 5c). 우점도지수(D)는 0.38~0.67의 범위로 그룹 1에서 가장 높고, 그룹 3에서 가장 낮게 나타났다(Fig. 5d).

Table 2.
Species composition of fish collected by bottom trawl in the southwestern sea of Jeju Island in summer 2020. N and W represent the number of individuals and biomass, respectively
3.5 그룹별 우점종 체장조성
각 그룹 해역의 우점 어종별 성숙개체 분포를 평가하기 위해 과거문헌을 통해 평가한 어종별 군성숙 기준 체장(L50)과 본 연구에서 조사한 어종별 체장빈도를 비교한 결과는 Figs. 6, 7, 8과 같다. 우점종으로 구분된 어종 중 군성숙체장이 연구되지 않은 꼼치, 두드럭게와 그룹 1에서 1개체만 어획된 넙치를 제외한 9종을 대상으로 성숙개체의 분포를 비교하였다.
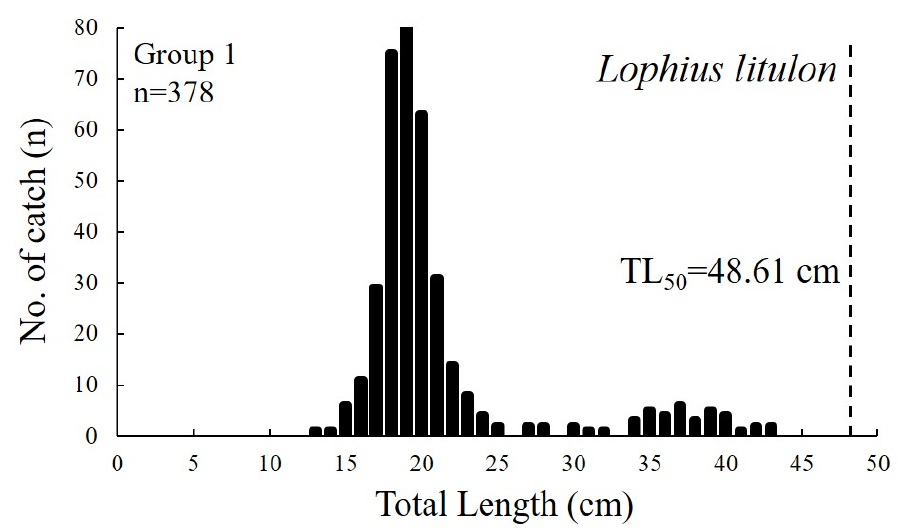
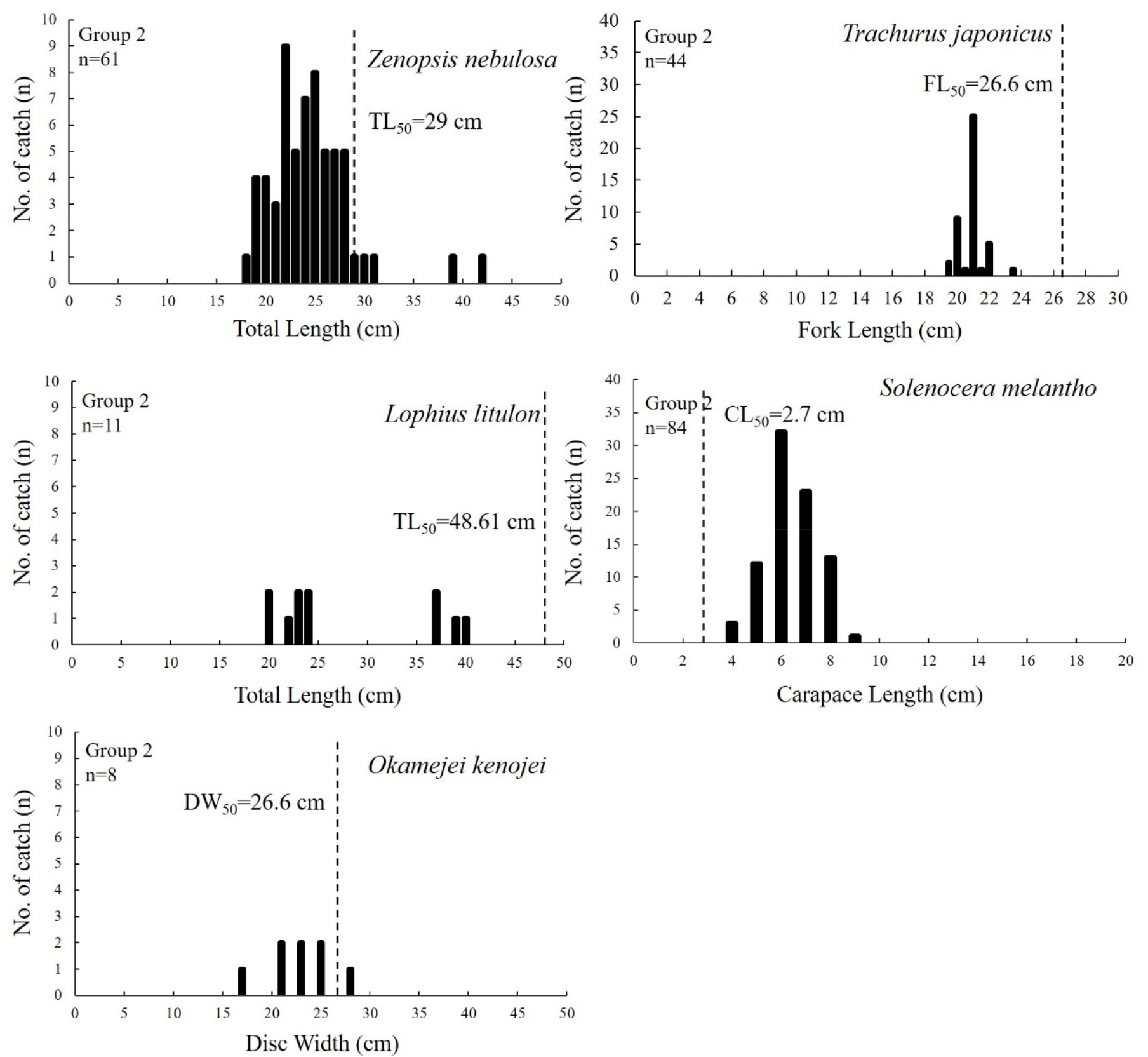
그룹 1에서 가장 우점하는 황아귀의 출현 전장(TL, Total Length)은 13~42 cm이며, 평균 전장은 21.01 cm로 나타났다. 군성숙전장 48.67 cm (Kim et al., 2020)과 비교시 모두 미숙한 개체가 분포하는 것으로 나타났다(Fig. 6). 그룹 2에서 가장 우점하는 민달고기의 출현 전장 분포는 18~42 cm, 평균전장은 24.5 cm로 나타났으며, 군성숙전장 29 cm (Froese and Pauly, 2020)과 비교시 모두 미숙한 개체가 분포하는 것으로 나타났다(Fig. 7). 다음으로 어획량이 높은 전갱이(Cha et al., 2009)와 황아귀(Kim et al., 2020)는 모두 미숙한 개체가 분포하는 것으로 나타났다. 대롱수염새우의 출현 갑각장(CL, Carapace Length) 분포는 3~9 cm, 평균갑각장은 6.3 cm (Oh et al., 2005)로 나타나 모두 군성숙 갑각장 이상의 개체가 분포하는 것으로 나타났다. 홍어의 출현 체반폭(DW, Disc Width)의 출현 범위는 17~28 cm, 평균체반폭은 22.9 cm로 나타났으며, 군성숙 체반폭 26.6 cm (Youn et al., 2022)과 비교시 성숙가능 개체는 1개체(0.5%)로 대부분 미숙한 개체가 분포하는 것으로 나타났다. 그룹 3에서 가장 우점하는 황아귀의 출현 전장 범위는 18~43 cm, 평균전장은 26.1 cm로 나타나 모두 미숙 개체가 분포하는 것으로 나타났다(Fig. 8). 달고기의 출현 전장 범위는 17~45 cm, 평균전장은 23.08 cm으로 나타났으며, 군성숙전장 이상(32.6 cm, Yoneda et al., 2006)의 개체는 1개체가 어획되어 대부분 군성숙전장 이하의 개체가 분포하는 것으로 나타났다. 붉은메기, 창꼴뚜기와 살오징어는 각각 어종별 전체 어획 개체수의 57% (n=4), 35.7% (n=10), 54.55% (n=6)의 비율로 군성숙 체장과 외투장(ML, Mentle Length) 이상의 개체가 어획된 것으로 나타나 다른 어종에 비해 상대적으로 성숙이 가능한 개체의 비율이 높은 것으로 나타났다.

4. 고 찰
본 연구에서 수행한 저층 해양생물 종조성 조사를 통해 여름 제주도 남서부해역에서 어획된 종 수는 총 37종으로 조사되었으며, 어류가 27종으로 가장 많이 어획되었다. 이 중 전체 어획량의 50% 이상을 차지하는 황아귀가 제주도 남서부 저층 해역에서 가장 우점하는 것으로 평가할 수 있다.
어종 별 생물량을 기반으로 한 대응분석 결과를 통해 제주도 남서부 저층 해역은 제주도 서부해역에서 동중국해 북부 외해로 이어지는 해역(그룹 1)과 제주도 남서부 근해 해역(그룹 2), 제주도 남부 외해(그룹 3)로 구분되었다(Fig. 3). Table 1과 Table 3을 통해 세 해역은 종 구성에 차이를 나타낼 뿐만 아니라 최대 수심에 차이를 나타냄을 확인할 수 있다. 그룹 1 해역은 수심 약 91 m 이내로 가장 얕고 황아귀, 꼼치가 우점하는 해역으로 평가되었으며, 그룹 2 해역은 수심 95 m 내외로 민달고기, 전갱이, 대롱수염새우가 우점하는 것으로 구분할 수 있다. 그룹 3 해역은 수심 약 107.5 m 내외로 황아귀, 달고기, 창꼴두기가 우점하는 것으로 나타나 그룹별 최대 수심과 종 구성에 차이가 있음을 확인 할 수 있다. 수심이 가장 얕은 그룹 1해역은 다른 해역과 비교하여 종 풍부도와 우점도가 높고 다양성 지수와 균등도 지수가 가장 낮게 나타났다. 그룹 2 해역은 그룹 1 해역과 비교하여 풍부도와 우점도 지수는 낮지만 다양성 지수와 균등도 지수가 높게 나타났으며, 그룹 3 해역은 가장 높은 종 다양성 지수와 균등도 지수, 가장 낮은 풍부도 지수와 우점도 지수를 나타내 그룹 1과 그룹 2 해역에 비해 총 어획 생물량은 가장 낮지만 가장 다양한 생물이 함께 분포하는 것으로 평가할 수 있다.
종별 생물량 구성이 비슷한 해역으로 구분된 그룹 1 해역의 정점별(St) 어종의 출현 종수와 생물량은 Fig. 9와 같다. St1은 St3과 St5에 비해 어획된 총 개체수는 3.3~7.2배, 총 생물량은 2~6.5배, 높은 것으로 나타나 제주도 서부 외해로 구분되는 St3와 St5보다 연안에 인접한 St1의 해양생물자원의 생물량이 높은 것으로 구분할 수 있다. 그룹 1의 정점(St)중 생물량이 가장 높게 나타난 St1의 어획 개체수와 생물량을 그룹 2, 그룹 3의 정점(St)과 비교하면 St1이 다른 두 그룹 보다 개체수는 1.3~1.8배, 생물량은 2~2.7배 높은 것으로 나타났다. 생태학지수 평가결과(Fig. 5)와 정점별(St) 조사 수심을 통해 종 다양성지수(H’)과 균등도지수(J)는 수심과 비례 관계, 종 풍부도지수(R)와 우점도지수(D)는 수심과 반비례 관계를 나타내어 수심이 얕을수록 종 다양성은 감소하고 특정 어종에 대한 우점도가 증가하는 것으로 해석할 수 있다. 따라서, 본 연구를 통해 제주도 남서부 해역의 저층 해양생물상은 외해(≥100 m) 보다 수심 90 m내외의 연안이 상대적인 종 다양성은 낮지만 총 생물량은 더 높은 것으로 평가할 수 있다. 또한, 그룹별 생물량 비중(Fig. 4b, Table 3)과 Fig. 5를 통해 여름 제주도 남서부 연안의 총 생물량 변동은 외해보다 특정 우점종의 생물량 변동에 더 큰 영향을 받으며, 특히, 여름 제주도 남서부 연안의 총 생물량은 가장 우점하는 황아귀의 생물량 변동과 관계가 있는 것으로 평가할 수 있다.
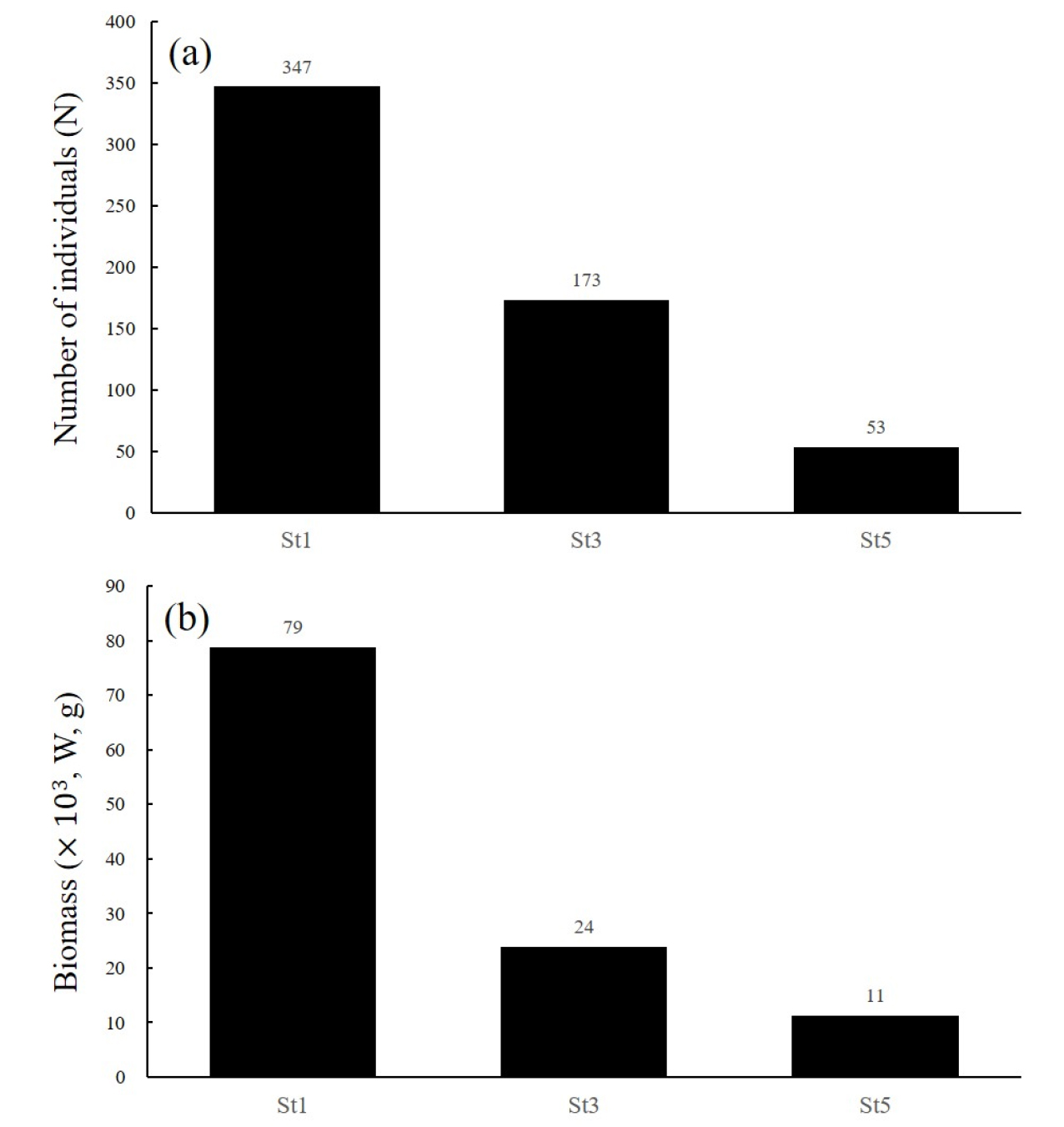
Table 3.
The dominant species in each group identified by correspondence analysis (CA) based on catch biomass by species
Kallianiotis et al.(2000)은 대륙붕과 대륙사면에서의 해양생물상의 변화는 수심 변화에 가장 큰 영향을 받는 것으로 평가하였으며, Smale et al.(1743)은 수심별 수온과 용존산소의 차이가 해양저서 생물군의 종 구성 변동과 관계가 있는 것으로 평가하였다. Lee et al.(2018)는 특정 해역의 수심별 어류 군집 종조성 차이는 수직적 수온 구조 차이와 관련 있는 것으로 평가하였으며, Huh and Kwak(1748)은 어류 생물량 변화와 해당 해역의 수온 변동은 양의 상관관계를 가지는 것으로 평가하였다. 또한, McConnaughey and Smith(2000)는 저어류의 공간 분포는 해저면의 퇴적물 타입과 관계가 있는 것으로 평가하였다. 따라서, 본 연구에서는 나타난 그룹별, 정점별(St) 우점종 구성과 생물량 차이는 조사 정점별(St) 수심 차이에 의한 수온 구조와 해저면의 환경과 관계가 있음을 예상할 수 있다.
정점별(St) 제주도 서남부 저층 해역에서 우점하는 해양생물 자원 중 황아귀(n=389)와 전갱이(n=44)는 모두 미숙한 개체가 나타났다. 북동중국해에 분포하는 황아귀는 여름(8~9월) 38°N 이북 해역까지 북상한 후 가을과 겨울에 산란을 위해 제주도 서남부 근해로 산란회유를 하는 것으로 알려져 있으며, 전갱이는 여름에 성장을 위한 섭이회류를 위해 양자강 주변의 중국측 연안(수심 <50 m)에 주요 분포 해역이 형성되는 것으로 알려져 있다(Park et al., 2000; Yoneda et al.,2002; Sassa et al., 2009). 어류 회유 범위와 분포 범위는 몸의 크기와 양의 상관관계로 몸의 크기가 클수록 회유 범위와 주요 분포 공간의 크기가 넓어 지는 것으로 알려져 있다(Nøttestad et al., 1749; Woolnough et al., 2009). 또한, 제주도 남서부해역에 분포하는 어류는 동중국해와 황해 사이를 계절에 따라 회유하는 특성을 가지고 있으며, 봄에 제주도 주변해역과 중국측 연안역에서 산란을 하고 여름에 섭이를 위한 북쪽으로 회유를 한 후 겨울에 다시 남하하는 특성을 나타낸다(Kim et al., 2007b). 따라서, 조사 시점에서 어획 되지 않은 황아귀와 전갱이 성어 개체는 어종별 회유 특성으로 인한 주요 분포해역의 북상과 연안으로의 이동으로 나타난 현상으로 예상할 수 있다. 어획 개체가 모두 성어로 구분되는 대롱수염새우(n=84)는 비교적 깊은 수심에 서식하며, 계절적으로 산란 또는 성장을 위해 회유하지 않고 특정 해역에 정착하여 서식하는 종으로 구분된다(Oh et al., 2005). 따라서, 제주도 남서부 해역의 대롱수염새우 개체군은 수심 95 m 내외의 연안(그룹 2)에 밀집하여 분포하고 있음을 확인할 수 있다. 또한, 그룹별 우점 종으로 구분되는 꼼치(n=22), 민달고기(n=61), 달고기(n=25), 붉은메기(n=9)는 성장에 따른 회유특성에 대한 연구 결과가 없거나 어획 개체수가 적으며, 해양환경 요인 중 수온에 가장 민감하게 반응하고 연중 산란(Kim et al., 2019)하는 두족류의 상대적으로 높은 성어 비율을 설명하기 위한 해양물리자료를 반영하지 못해 생물학적 특성에 기반하여 Figs. 7, 8에 나타난 어종별 성어와 미성어의 비율 차이를 설명하기에 어려움이 있다. 따라서, 해당종들의 공간분포 특성 변화를 파악하기 위해 어종별 생물학적 특성에 대한 추가 연구와 해양물리적 특성을 함께 고려한 연구가 필요하다.
본 연구는 여름 저층 생물종 조성에 한정하여 연구를 수행하였기 때문에 계절별, 수온환경 변동에 따른 종 구성 변화를 해석하기에 한계를 가지고 있다. 과거연구를 통해 제주도 주변해역을 포함한 동중국해의 수심별 수온은 모든 수층에서 장기적으로 상승하는 경향을 나타냈으며(Seong et al., 2010; Kwak et al., 2025), 이러한 수온 상승은 해당해역의 어류 공간분포에 영향을 미치는 것으로 연구되었다(Perry et al., 2005; Jung et al., 2014). 따라서, 장기적인 수온상승에 대응하여 제주도 남서부해역의 전반적인 어류 생물상 변화를 파악하기 위해 동일해역을 대상으로 계절별 수온 변동과 종 구성 변화를 함께 해석하기 위한 지속적인 조사와 종 구성 변화 평가 연구가 필요하다. 또한, 제주도 남서부해역은 대마 난류의 유입량 변화에 따라 생태계 구조가 변동하고 한국, 일본, 중국에서 경제적으로 중요한 온대성 표층 부어류가 중심을 이루는 해역이다(Kim et al., 2007b; Yukami et al. 2009). 따라서, 본 연구에서 평가한 저층 해양 어류 종조성 결과를 표층과 중층의 생물상 조사 결과와 함께 해석한다면 북동중국해 해양생물상을 구체적으로 평가할 수 있을 뿐만 아니라 수산경제학적 해역의 가치를 함께 평가할 수 있을 것으로 기대한다.
