

1. 서론: 연구 배경
2. 연구의 의의
2.1 전 지구 기후변화 대응을 위한 북극 연구의 과학적 기여
2.2 국제적 규범 이행 및 해양‧수산자원 관리 기반 마련
2.3 국가 차원의 선제적 투자 및 국가 과학기술 역량 강화
2.4 국가 위상 제고 및 정책 기반 구축
3. 연구개요 및 내용
3.1 연구지역 및 탐사 개요
3.2 연구내용 및 구성
4. 주요 연구성과 및 결과
4.1 서북극해에서 대서양화 현상 최초 발견
4.2 해양환경 변화에 따른 식물플랑크톤 분포와 군집 구조의 장기 변동
4.3 식물플랑크톤 계절 주기와 일차 생산성 변화: 장기 계류 관측을 통한 분석
4.4 북극 대구(Boreogadus saida)의 분포 연구: eDNA 기법을 활용한 새로운 접근
4.5 서북극해에서 대서양 기원수 확장에 따른 생지화학 변화 현상 최초 규명
4.6 서북극해에서 급속한 해양 산성화와 생지화학적 조절 요인
4.7 북극 담수 유입 증가에 따른 생태계 반응 모델
4.8 퇴적물 트랩을 활용한 서북극해 탄소플럭스 변동 연구
5. 결론 및 향후 전망
1. 서론: 연구 배경
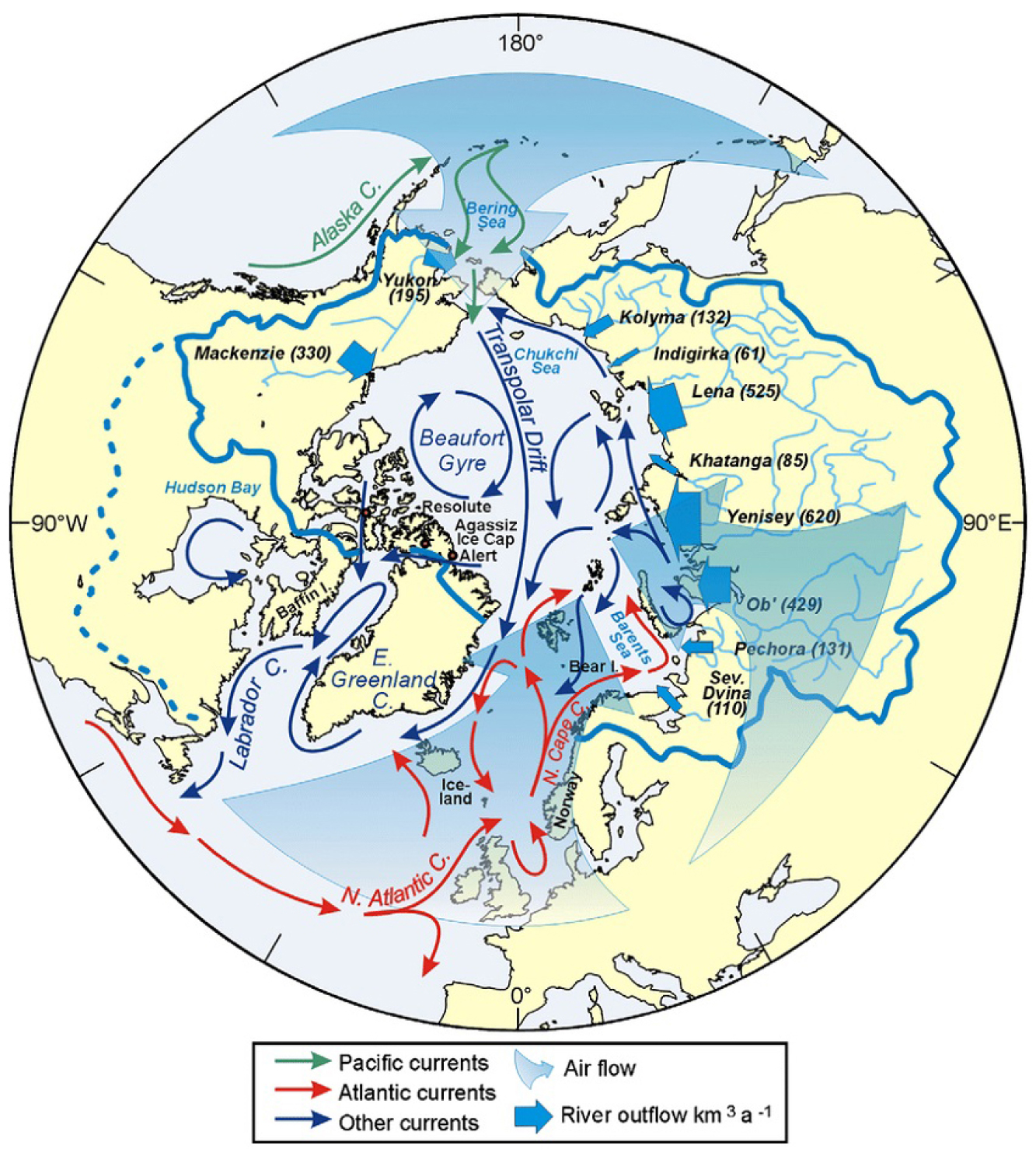
북극해는 유라시아 대륙, 북아메리카 대륙과 그린란드로 둘러싸인 전 세계 해양 면적의 약 2%를 차지하는 가장 작고 수심이 얕은 해양이지만, 지구 해양 순환과 기후 시스템에 결정적인 영향을 미치는 핵심 지역이다(Wang et al., 2016; Meredith and Garabato, 2021; Rantanen et al., 2022) 북극해는 경도 0°와 180°를 기준으로 서북극해(Western Arctic Ocean)와 동북극해(Eastern Arctic Ocean)로 구분된다. 서북극해는 베링 해협을 통해 유입되는 태평양 기원수(Pacific Water)의 영향을 받으며, 동북극해는 프람 해협과 바렌츠해를 통해 유입되는 대서양 기원수(Atlantic Water)의 영향을 받는다(Fig. 1) (Macdonald et al., 2005; Rudels and Carmack, 2022). 상대적으로 따뜻한 두 해수의 유입은 북극해의 열‧염분 구조, 생지화학적 순환, 생물다양성 그리고 전 지구 기후 변동성에 중요한 영향을 미친다(Ramudu et al., 2018; Ardyna and Arrigo, 2020; Jung et al., 2021; Alabia et al., 2023; Polyakov et al., 2023; Ye et al., 2024; Catherine et al., 2025).
최근 수십 년간 지구 온난화가 가속화되면서 북극은 전 지구에서 가장 빠르게 더워지고 있는 지역이 되었으며, 그 온난화 속도는 전 지구 평균 기온 상승의 4배 이상 빠르게 상승하고 있다(Rantanen et al., 2022). 이로 인해 북극해 해빙은 빠른 속도로 사라지고 있으며, 1979년 이후 북극해 해빙 면적은 9월 평균 약 42% 감소하였다(Fig. 2)(Moon et al., 2024). 최근 연구에서는 북극 여름 해빙이 완전히 소멸될 시점을 기존의 2050년에서 2030년대로 앞당겨 질것으로 예측하고 있으며(Kim et al., 2023), 이는 기존 기후모델이 북극의 복잡한 상호작용을 충분히 반영하지 못하고 있음을 시사한다.
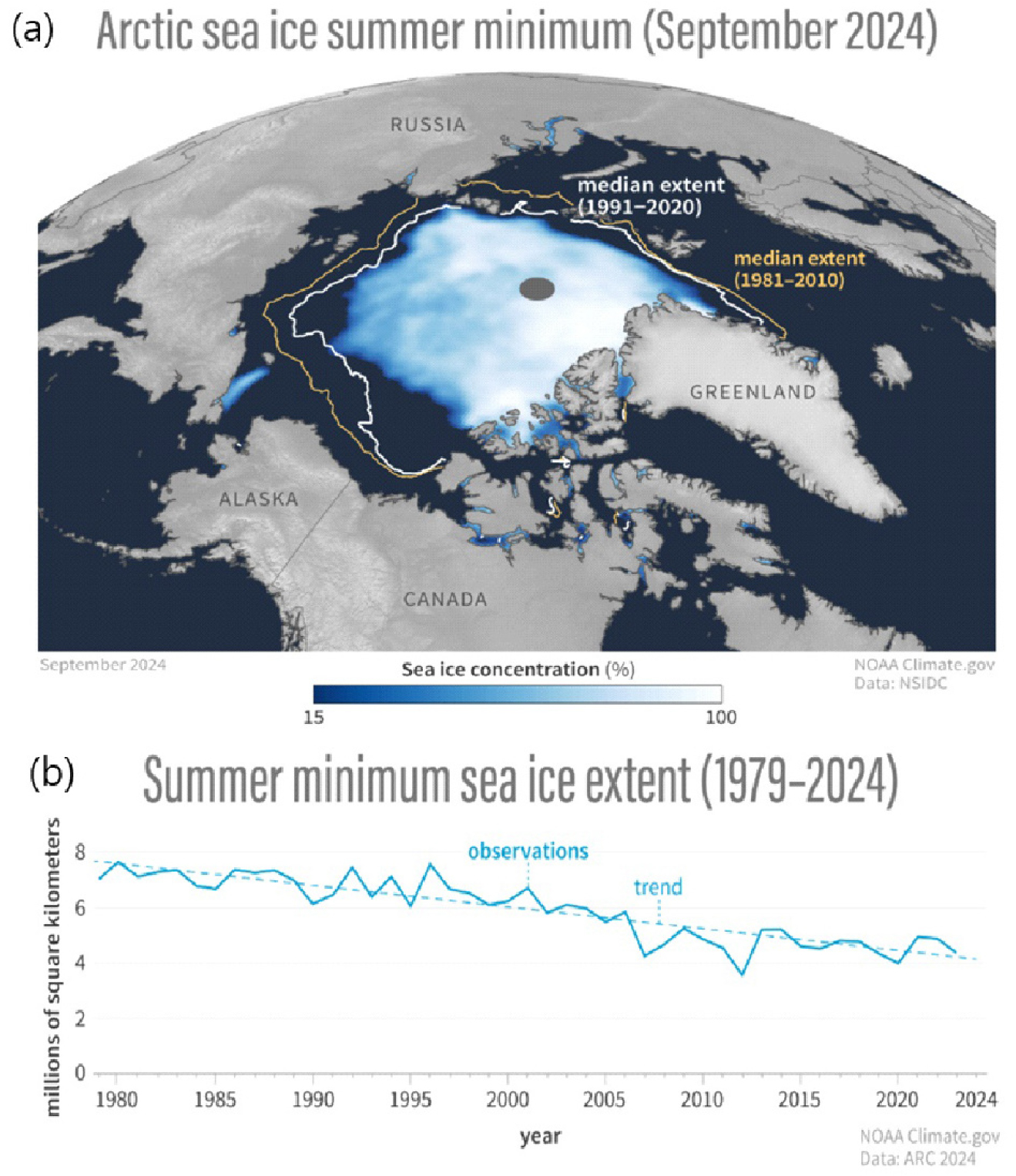
Fig. 2.
Arctic Sea ice summer minimum (a) and summer minimum sea ice extent (b) (Figure from ‘NOAA Climate.gov’ website (https://www.climate.gov) and Moon et al., 2024).
급격한 북극 온난화와 해빙 감소는 단순히 지역적 현상에 그치지 않고, 한반도를 포함한 중위도 지역에서 빈번하게 발생하는 한파, 폭염, 가뭄 등 극단기상 현상의 주요 원인으로 지목되고 있다. 북극권의 환경 변화는 해빙 감소, 해양 열 저장 증가, 대기 순환의 변화 등 여러 요소가 복합적으로 작용하는 기후 되먹임(climate feedback) 메커니즘에 의해 가속화되고 있으며, 이는 전 지구적인 기후 시스템의 불안정성을 증폭시키는 원인이 된다(Cohen et al., 2014; Kim et al., 2021a). 특히 해빙 감소는 북극해의 해양 물리‧화학적 구조를 근본적으로 변화시키고 있다. 서북극 보퍼트해(Beaufort Sea)에서는 지난 30년간 염분약층의 해양 열량이 약 2배 증가하였으며, 이는 해빙 면적 감소, 태양 복사열 증가, 그리고 베링해협을 통해 유입되는 따뜻한 태평양수의 온도 상승과 밀접한 관련이 있다(Timmermans et al., 2018).
북극해는 전 세계 하천 유량의 약 11% 이상을 차지할 만큼 담수 유입이 활발한 해역이다(Osborne et al., 2018; Meredith and Garabato, 2021). 강수 증가와 영구동토층 해빙(melting)으로 인한 하천 유량 증가는 북극해의 담수화를 가속화시키고 있으며, 이는 수온 및 염분 구조 변화, 해류 순환, 해빙 분포에 직접적인 영향을 미친다(Rudels and Carmack, 2022). 특히 북극해로 유입되는 육상 기원 용존유기탄소(DOC) 농도의 증가로 탄소 순환 및 산성화를 가속화하여 전 지구적 탄소 균형에도 영향을 미친다(Semiletov et al., 2016; Jeon et al., 2025). 특히 서북극해는 전세계 해양에서 산성화 속도가 4배까지 빠르게 진행되는 것으로 보고되었으며(Qi et al., 2017; Qi et al., 2022), 이 결과들은 북극 해양생태계의 기반 구조를 위협하고 있다.
최근 북극의 온난화 가속화는 북극해 생태계의 구조와 기능을 빠르게 변화시키고 있다. 북극해와 북태평양의 연간 총 어획량은 약 4,600만 톤으로 전 세계 수산물 생산량의 40%을 차지하지만, 해빙 감소와 수온 상승에 따라 2100년까지 약 40종 이상의 어류 서식지가 변동할 것으로 예측된다(Field et al., 2014; FAO, 2022). 난류성 어종이 북상하면서 북극은 새로운 어장으로 부상하고 있으며, 냉수성 어종은 점차 사라지고 있다(Cheung et al., 2010; Poloczansk et al., 2013). 특히 태평양 기원 동물플랑크톤(Eucalanus bungii 등)과 무척추동물 유생(Meroplankton)의 북상 현상이 뚜렷해지고 있다(Ershova et al., 2019; Kim et al., 2020). 이는 북극 생태계가 점차 고유종에서 아북극종으로 변화하고 있음을 보여주며, 북극 해양 먹이망 구조의 근본적인 변화를 의미한다. 이러한 급격한 북극해 생태계 변화 속에서 2021년 발효된‘중앙 북극 공해상 비규제 어업방지협정(Central Arctic Ocean Fisheries Agreement, CAOFA)’에 따라 북극 공해상에서 생물자원 변화에 대한 과학적 규명과 국제공동연구 수행은 단순한 학문적 선택이 아니라 국가적 책임이자 의무가 되었다.
본 연구는 북극 온난화에 따른 해양환경 및 생태계 변화를 종합적으로 규명하고, 물리‧화학‧생물학적 상호작용을 통합적으로 이해하는 것을 목적으로 한다. 이를 위해 쇄빙연구선 아라온(ARAON)을 이용한 장기적이고 지속적인 북극해 탐사를 수행하여, 급변하는 북극 해양 시스템의 특성과 그 변화 메커니즘을 심층적으로 이해하고자 한다. 이를 위해 해빙-해양 물질 교환, 해양의 물리적 특성, 탄소 순환, 플랑크톤 생태, 수산자원 변동성 등을 장기 관측과 모델링을 통해 연구를 수행하고 있다. 또한 국제 프로그램과의 연계를 강화하여 한국의 과학적 기여를 확대하고, 국제사회의 지속가능한 북극 관리 및 기후변화 대응에 기여하고자 한다. 본 종설 논문에서는 해양수산부의 재원으로 해양수산과학기술진흥원의 지원을 받아 수행중인‘북극해 온난화-해양생태계 변화 감시 및 미래전망 연구’과제를 소개하고, 이 연구 과제를 통해 밝혀진 연구 결과들에 대해 소개하고자 한다.
2. 연구의 의의
2.1 전 지구 기후변화 대응을 위한 북극 연구의 과학적 기여
북극은 지구 기후시스템의 핵심 조절자로서, 온도 상승과 해빙 감소 등과 같은 변화가 전 지구적인 기후 체계 변화로 이어질 수 있는 중요한 지역이다. 따라서 북극 온난화로 인한 물리적‧화학적‧생태학적 변화를 종합적으로 규명하고, 이를 통해 지구 기후시스템의 변동성을 이해하며 기후위기에 대한 선제적 대응 기반을 구축하는 것은 인류가 직면한 필수 과학적 과제이다. 본 연구는 서북극해에서 장기 관측과 통합 분석을 통해 북극 해빙-해양환경 및 생태계 변화를 정량적으로 진단하고, 이상변화 현상을 조기에 탐지할 수 있는 과학적 근거를 마련함으로써 전 지구 기후 안정성 회복과 국제사회의 기후변화 대응 노력의 실질적 이행에 기여하고자 한다.
2.2 국제적 규범 이행 및 해양‧수산자원 관리 기반 마련
한국을 포함한 10개국이 뜻을 모아 2021년 발효된 중앙북극 공해상 비규제 어업 방지 협정(CAOFA)에 따라 협정국들은 북극해에서 향후 16년간 과학적 연구만을 허용하고 상업적 조업을 제한함으로써, 과학적 조사와 생물자원 모니터링의 국제적 의무를 수행해야 한다. 북극해는 인류 공동의 자산이자 생물자원의 보고로, 해빙 감소와 수온 상승에 따라 북반구 어류의 북상과 신규 어장 형성이 가속화되고 있다. 이러한 변화는 자원 관리, 생태계 보전, 그리고 지속가능한 이용을 위한 과학적 근거 구축을 필수적으로 요구한다. 본 연구는 CAOFA의 이행을 지원하고, 북극 수산자원과 생태계 변화를 정량적으로 진단하여 국제 공동연구와 정책 협력의 기반을 마련하고자 한다. 이를 통해 북극 공해상 어족자원의 보호 및 지속가능한 조업권 확보를 위한 과학적 근거를 제시하고, 한국이 국제 북극 정책 논의와 공동과학연구에서 주도적 역할을 강화하는 데 기여할 것이다.
2.3 국가 차원의 선제적 투자 및 국가 과학기술 역량 강화
북극은 지구 기후변화의 최전선이자 새로운 과학적‧경제적 기회의 공간으로, 이를 이해하고 대응하기 위하여 정부 차원의 지속적 투자와 과학적 연구가 필수적이다. 한국은 이미 쇄빙연구선 아라온(ARAON), 북극해 장기모니터링 시스템(Korea Arctic Monitoring System, KAMS), 북극 해양환경 데이터시스템(Korea Arctic Ocean-Data System, KAOS, https://kaos.kopri.re.kr) 등 세계적 수준의 연구 인프라를 구축하고 있다. 이러한 기반을 활용해 장기 관측과 정밀 분석을 수행하면서, 북극권 국가들과의 협력연구를 확대하고 국제 공동프로그램(Pacific Arctic Group, PAG; Distributed Biological Observatory, DBO; Synoptic Arctic Survey, SAS; CAOFA 등)에서 주도적 역할을 수행할 수 있다. 또한, 국가 차원의 지원은 북극 해양탐사 및 기초자료 확보뿐만 아니라, 극지 과학기술 인력 양성, 해빙-해양-생태계 통합 모델링 등 국가 과학 역량의 구조적 강화로 이어진다. 이는 단순히 연구 성과 축적을 넘어, 향후 북극 연구의 패러다임을 선도하는 핵심 동력이 될 것이다.
2.4 국가 위상 제고 및 정책 기반 구축
북극 해빙 소멸과 해양환경 변화에 대한 정밀 관측과 예측 연구는 단순한 학문적 성과를 넘어, 기후 변화 대응과 지속가능한 해양 관리 전략 수립을 위한 핵심 과학 자산이다. 신뢰도 높은 관측 및 예측자료는 북극항로 운영, 해양 안전 및 재난 대응, 한반도 기후 예측, 극지 자원 탐사 등 다양한 분야의 과학적 의사결정에 활용될 수 있으며, 국가의 기후 리스크 관리 역량을 강화하는 기반이 된다. 본 연구를 통해 확보된 자료는 북극 해양환경 데이터 시스템(KAOS)을 통해 관리‧공개되며, 이는 국가 정책 수립, 북극항로 안전망 구축, CAOFA 대응, 이상기후 대응 등에 기여함으로써 한국의 북극 연구 리더십을 강화할 것이다.
3. 연구개요 및 내용
3.1 연구지역 및 탐사 개요
본과제는 북태평양수의 영향을 직접받는 베링해협과 척치해 그리고 동시베리아해역으로 구성된 서북극해(western Arctic ocean)에서 연구를 수행하였다(Fig. 3). 특히 동시베리아 북부 해역은 대서양화로 인한 북극해 환경변화를 탐지하고 변동성을 예측하는 데 매우 중요한 미답해역으로, 보고된 자료가 드물어 희소성이 높은 자료가 생산되는 해역이다. 현재 동시베리아해는 우리나라와 미국 NABOS (Nansen and Amundsen Basins Observational System) 팀이 공동으로 연구를 수행하고 있다. 또한 베링해협에서 수행하고 있는 DBO 3 관측 라인은 pan-Arctic DBO 프로그램의 일환으로 수행되고 있다. 본 연구는 쇄빙연구선 아라온을 활용하여 2016년부터 매년 7월~8월에 서북극해에서 탐사를 수행하였다.
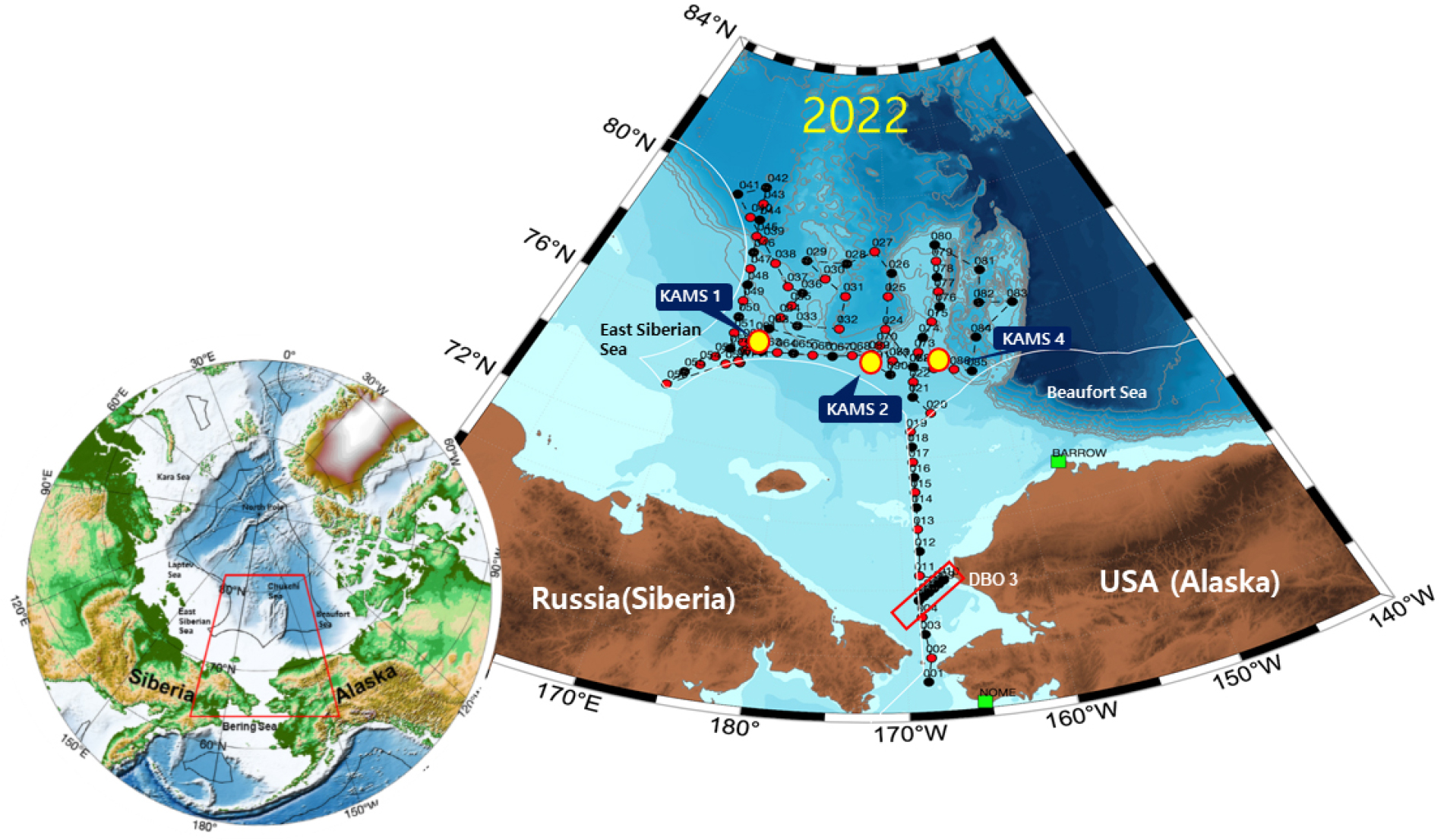
Fig. 3.
Map showing the East Siberian Sea, Chukchi Sea, and Bering Strait which are the main study areas surveyed using the IBRV Araon. The surveys in the East Siberian Sea and Chukchi Sea are limited to international waters. The White line indicates the Exclusive Economic Zone (EEZ). KAMS means Korea Arctic Mooring System (Figure created by K.H. Cho).
3.2 연구내용 및 구성
본 연구는 북극해 온난화로 인한 해양환경과 생태계 변화를 종합적으로 규명하고 미래 변동을 전망하기 위해 설계되었다. 연구는 네 개의 핵심 축으로 구성된다. 첫째, 서북극해 해양-해빙 현장탐사 및 온난화 감시망 구축을 목표로 하고 있다. 해양환경 모니터링을 위한 ‘북극해 장기모니터링 시스템(KAMS)’ 3기와 웹기반 ‘북극해양환경 데이터 시스템(KAOS)’를 운용하면서 국내외 연구자들에게 북극해 자료를 공유한다. 둘째, 해양-해빙간 물리적 변동성과 물질순환 과정 연구를 통해 수온, 염분, 담수유입, 영양염, 이산화탄소, 미량금속 및 미세플라스틱의 변화를 정량적으로 파악하고, 환경변화가 탄소순환 및 해양 산성화에 미치는 영향을 규명한다. 셋째, 해양 생태계 및 생물수산자원 반응 연구를 통해 해빙 미세조류, 플랑크톤, 어류 등의 시‧공간적 분포 특성과 먹이망 구조를 분석하며, 환경 DNA를 활용한 새로운 자원 모니터링 체계를 확립한다. 넷째, 인간활동 시나리오에 따른 북극해 환경변화 미래전망 연구를 수행하기 위하여 대기-해양 간 에너지 교환 과정과 북극해 수문‧생지화학 순환을 개선한 지구시스템 모델을 구축하고, 이를 바탕으로 다중 시나리오 기반의 북극해 미래 환경변화 예측체계를 구축한다. 이러한 연구는 현장관측, 장기계류, 위성자료, 모델링 및 KAOS 와 연계하여 추진되며, 궁극적으로 북극해의 급격한 환경변화에 대한 조기진단, 과학적 예측, 그리고 국제사회가 요구하는 기후‧생태계 대응 기반 마련에 동참하고자 한다.
본 연구과제는 국내에서 수행되고 있는 유일한 북극해 종합 해양연구로서 현재 7개 연구기관(경상대, 부경대, 부산대, 전북대, 포항공대, 한양대, 한국해양과학기술원)과 2개 산업체(네오씨텍, 선진해양)가 합류하여 연구를 수행하고 있다. 또한 국제 협력 파트너로는 유럽 연합의 Arctic PASSION (Pan-Arctic Observing System of System: Implementing Observations for Societal Needs) 프로젝트, 미국 알라스카 대학의 NABOS 프로젝트, 일본의 ArcS II (Arctic Challenge for Sustainability II) 프로젝트 및 미국에서 주도하는 DBO 프로그램과 국제공동 연구를 수행하고 있다.
4. 주요 연구성과 및 결과
4.1 서북극해에서 대서양화 현상 최초 발견
북극해에서 대서양화는 대서양 기원의 따듯하고 염분이 높은 수괴가 북극해로 유입되어 확장되는 현상을 의미한다. 이는 지난 수십 년간 해빙의 급격한 감소와 북극 대기 및 해양의 온난화를 나타내는 고위도 기후 변화의 중요한 요소로서 고려되었다(Polyakov et al., 2020; Wang et al., 2025). 최근 연구에 의하면, 서북극해는 빠르게 진행 중인 대서양화의 새로운 중심지로 부상하고 있다. 기존에는 주로 동북극해(Barents-Kara-Laptev Sea)에서 대서양 기원수의 온도 상승과 상부 확장(shoaling)이 보고되었으나, 최근 20여 년간의 장기 관측 결과 서북극해인 동시베리아해에서 대서양화의 증거를 최초로 보고하였다(Polyakov et al., 2025). Polyakov et al.(2025)는 동시베리아 해역(East Siberian Sea)과 마카로프 분지(Makarov Basin)에서 20년 이상의 계류관측를 분석하여, 대서양 기원수 상부 경계면의 수심이 약 160 m에서 65 m로 상승하였음을 관찰하였다(Fig. 4). 이로 인해 대서양 기원의 따뜻하고 염분이 높은 수괴가 상층 혼합층과 직접 접촉하면서, 겨울철 환기(winter ventilation)가 활발해지고 상향 열플럭스가 증가하여 해빙의 계절적 소멸을 촉진시키는 것으로 나타났다. 이러한 해양 열수송 증가는 북극해의 해빙-대기 간 열교환 구조를 변화시키고, 해빙 두께 손실(0.7~1.0 m) 및 여름철 해빙 소멸 시기가 앞당김을 유발하는 주요 요인으로 작용하고 있음을 밝혔다. 특히 본 연구는 대서양화가 로모노소프 능선을 넘어 마카로프 분지로 확장되고 있음을 처음으로 실측으로 제시하였다(Polyakov et al., 2025). 또한 2015~2023년 여름철 현장관측을 통해 염분층 약화, 영양염 상향이동, 엽록소 최대층의 상부확장등이 확인되었으며, 이는 북극 생태계 구조와 생산성에 직접적인 영향을 미치는 초기 생태계 반응임을 입증하였다(Polyakov et al., 2025). 따라서 대서양화의 확장은 북극해 대기 및 해양 순환 변동, 기상‧해양 재해, 해양생태계 변화에 직‧간접적인 영향을 줄 수 있으므로, 북극해의 변동성을 안정적이고 지속적으로 파악하기 위해서는 장기 해빙-해양 계류장비와 지속적인 현장자료 확보가 필요하다.
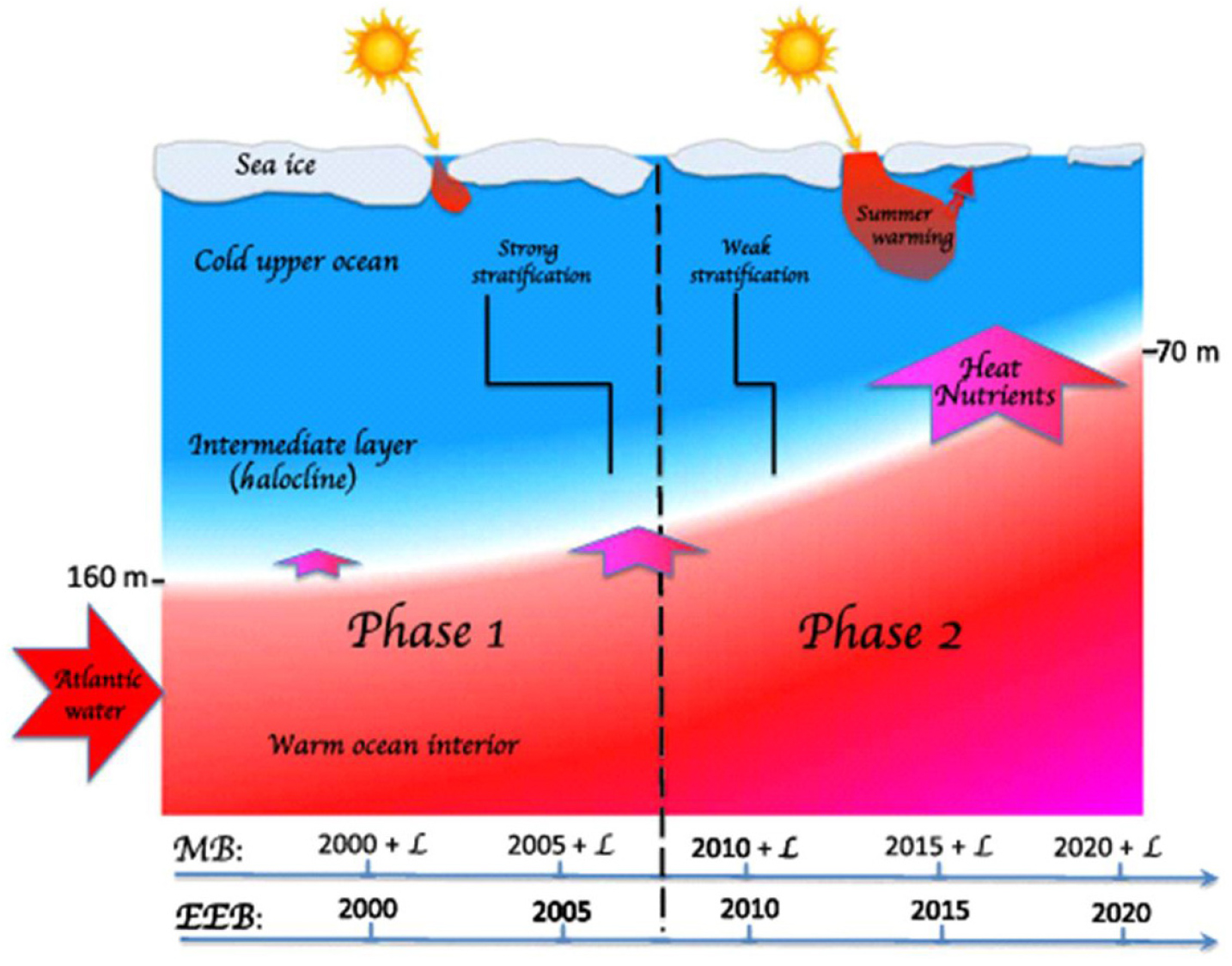
Fig. 4.
Two phases of atlantificationof the Arctic Ocean. Phase 1: warming of the ocean interior and AW/nitriclineshoaling. The halocline remains relatively strong, preventing deep ventilation. Phase 2: halocline weakened enough to allow deep ventilation. The shoaling of Atlantic water from ~150 m in the early 2000s to up to 70 m in recent years in the eastern Eurasian Basin has coincided with the seasonal disappearance of the halocline, which enhances upward heat and nutrient transport from the ocean interior to the upper ocean, causing fast sea-ice Loss and an increase in primary production (Figure from Polyakov et al., 2025).
4.2 해양환경 변화에 따른 식물플랑크톤 분포와 군집 구조의 장기 변동
북극해 식물플랑크톤은 북극 해양 생태계의 기초 생산자이자 먹이사슬의 근원으로서 해양생태계에 핵심적인 역할을 한다. 최근 북극 해빙의 지속적인 감소로 인한 일사량 증가와 성층 강화로 북극해의 일차생산은 1998년 대비 2018년에 약 57% 증가하면서 큰 변화를 겪고 있다(Lewis et al., 2020). 서북극해에서 식물플랑크톤의 분포 및 군집특성을 살펴보기 위하여 척치해 및 동시베리아해에서 해양 환경 변화가 식물플랑크톤 생체량과 군집 구조에 미치는 영향을 파악하기 위하여 다년간 자료를 분석하였다(Lee et al., 2023; Lee et al., 2025). 해당 지역은 뚜렷하게 동시베리아와 척치해에서 환경적으로 지역적 차이를 보였다. 동시베리아해는 저온‧고염수와 높은 영양염, 높은 해빙농도가 특징적이었고, 척치해는 따뜻하고 성층화가 강한 빈영양 환경이 특징적이었다(Figs. 5 and 6). 2017년 이후 대서양 기원수가 동시베리아해 북부로 확장되면서 약 30 - 40 m 깊이에 위치한 영양염약층(nitracline)이 얕아지고, 표층으로의 질산염 공급이 증가하였다. 이러한 변화는 식물플랑크톤 성장의 주요 제한 요소였던 영양염 가용성을 개선하였으며, 일조량이 풍부했던 2018년에는 수층 엽록소-a 생체량이 83.73 mg m-2로 최대치를 기록하였다(Fig. 5). 이 시기에는 규조류(diatoms)가 우점하며 대형 식물플랑크톤 중심의 생태계가 구성되었다. 반면 2019~2020년에는 해빙의 빠른 후퇴와 수온 상승에도 불구하고 일조량 감소와 강한 성층화로 인해 생체량이 전반적으로 감소하였고, Prymnesiophytes와 같은 소형 편모조류(flagellates)가 우점하는 생태계로 전환되었다(Fig. 7). 이러한 군집 구조의 변화는 북극 생지화학적 순환 및 탄소 플럭스에 중대한 영향을 미치는 신호로 해석된다. 규조류에서 소형 편모류로의 전환은 생물학적 탄소펌프의 효율을 약화시키며, 탄소가 표층에서 재순환되는 미세생물 루프(microbial loop) 중심의 먹이망으로 변화시킨다. 따라서 북극해의 탄소순환 및 생태계 변화를 이해하기 위해서는 해빙, 영양염, 성층화, 일조량 등 복합적 요인을 통합적으로 관측하는 장기 모니터링이 매우 중요하며, 향후 북극 생태계의 생산성 변화 예측과 탄소순환 모델 고도화에 필수적인 과학적 근거를 제공한다.
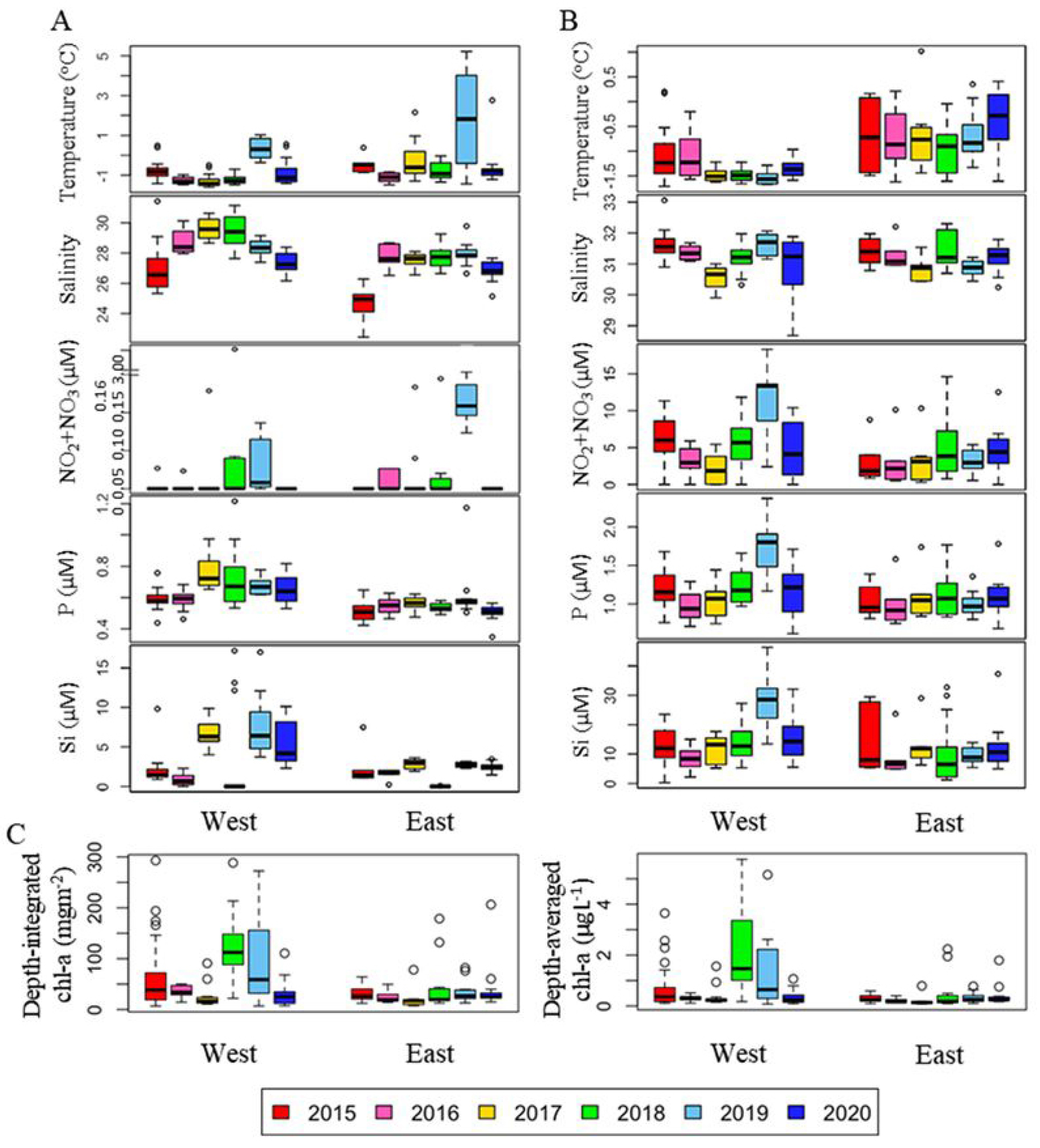
Fig. 5.
Annual variations in physical, chemical and biological variables in the (A) surface and (B) subsurface chlorophyll maximum (SCM) layers. The box plot represents the data on the east and west sides during six cruises from 2015-2020. The central mark in the box is the median, and the edges of the box are the 25th and 75th marks. NO2+NO3, nitrite and nitrate; P, phosphate; Si, silicate (Figure from Lee et al., 2023).
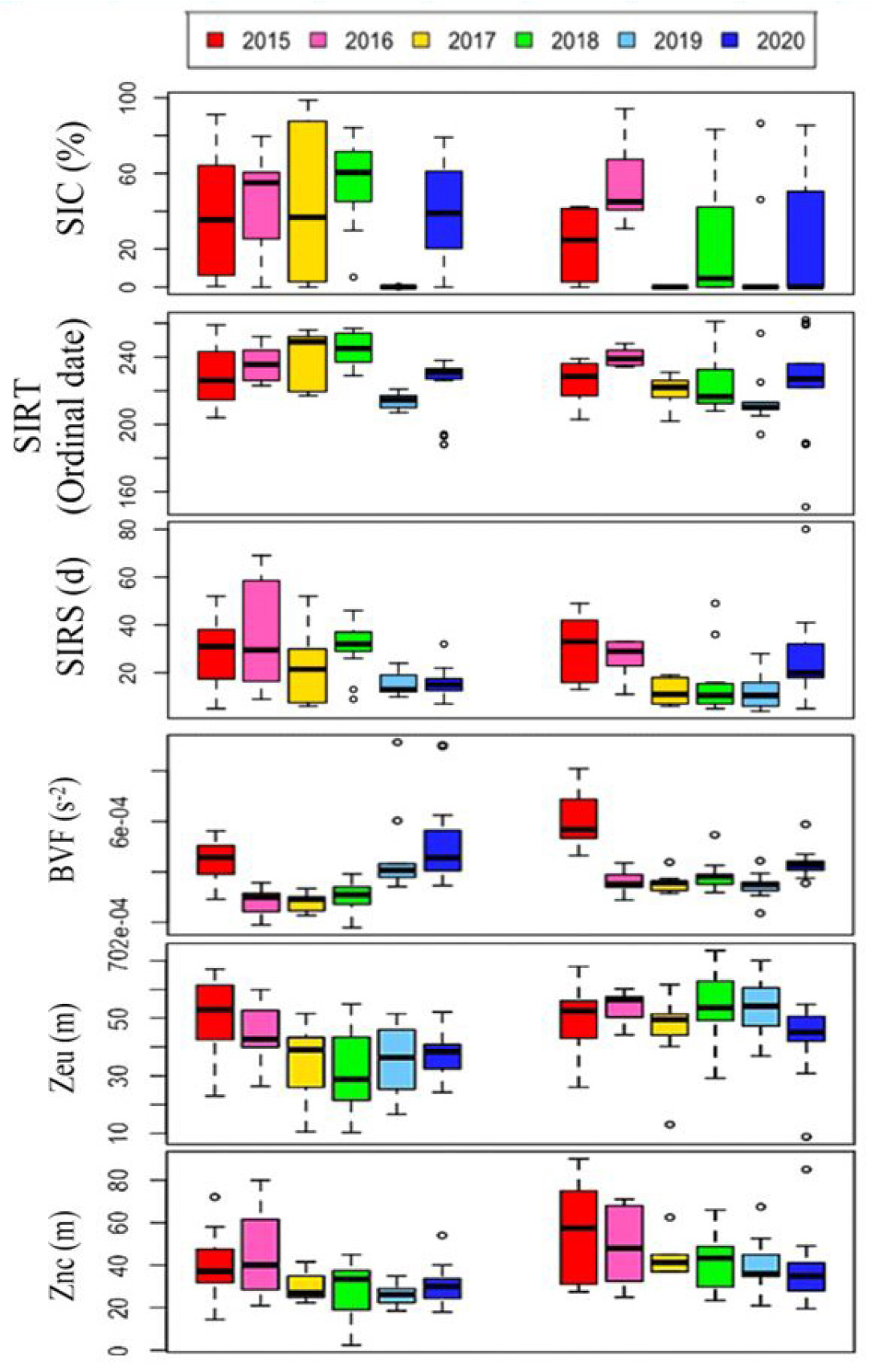
Fig. 6.
Annual variations of 7-day average sea ice concentration prior to sampling period, sea ice retreat timing (SIRT), and sea ice retreat speed (SIRS), BVF, mean Brunt-Väisäläbuoyancy frequency (N2, s-2) upper 100 m depth; Zeu, euphotic depth (m); Znc, nitracline depth (m) in the east and the west sides during the cruises (This figure is modified from Lee et al., 2023).
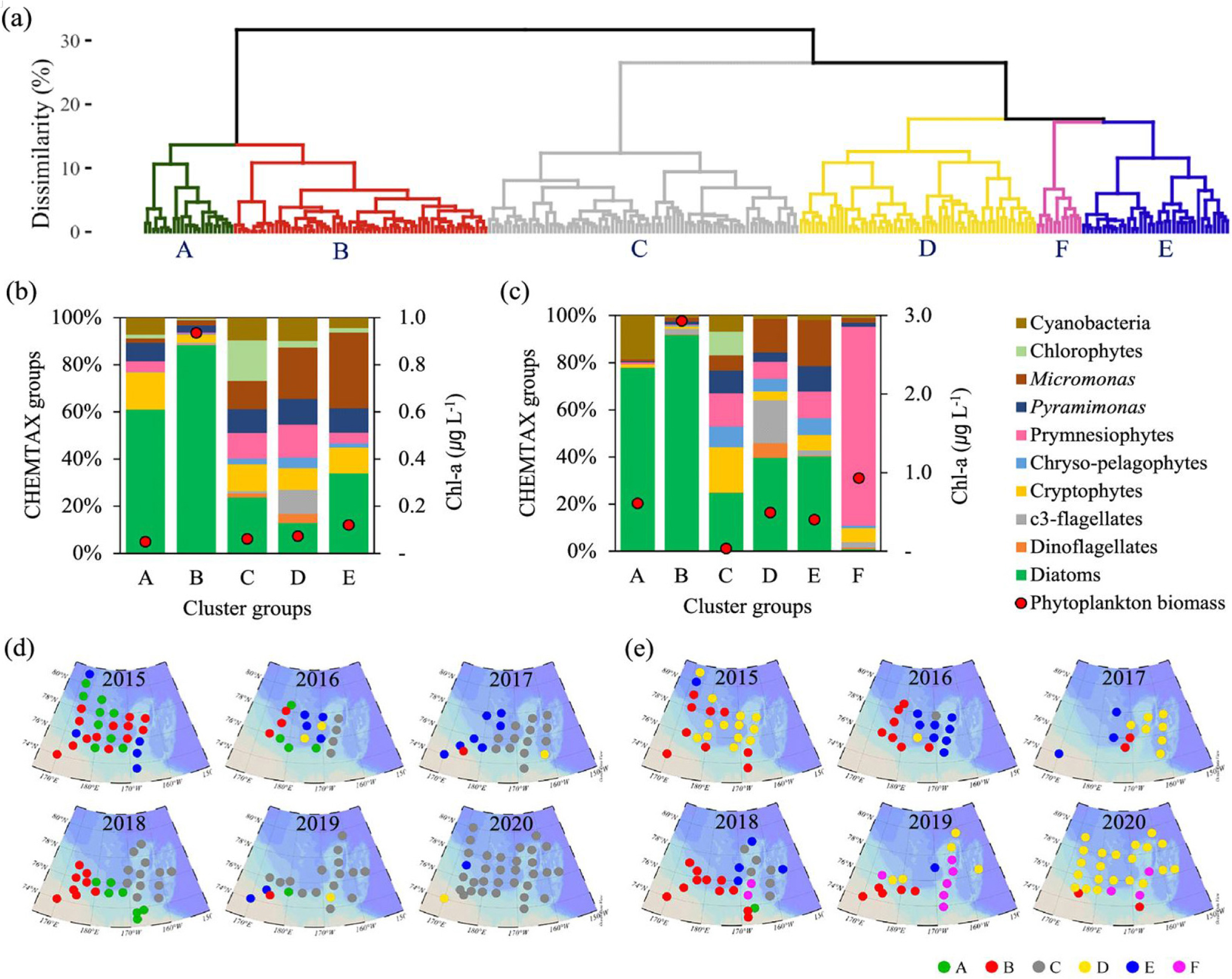
Fig. 7.
Classification of the samples in (a) a dissimilarity dendrogram based on phytoplankton group chl-a contribution at each station and depth. The average chl-a concentrations and the proportion of chl-a contributed by each phytoplankton group for each cluster in (b) the surface and (c) the subsurface chlorophyll maximum (SCM) layers of the study area. Comparison of the spatial distribution of phytoplankton communities (cluster groups) in (d) the surface and (e) the SCM layers (Figure from Lee et al., 2025).
4.3 식물플랑크톤 계절 주기와 일차 생산성 변화: 장기 계류 관측을 통한 분석
접근이 어려운 고위도 동시베리아해 및 척치해 북부에서 식물플랑크톤의 계절 주기와 일차 생산성 변화를 규명하기 위해 KAMS에 설치된 Chl-a 형광계를 이용하여 장기 모니터링 자료를 분석하였다(Ko et al., 2024; Park et al., 2025). 해빙 조류들은 해빙이 여전히 높은 농도(>90%)를 유지하는 5~6월에 이미 해빙하 대번성(Under Ice Blooms, UIBs)이 시작되어, 위성으로 관측 가능한 해빙 후퇴 시기보다 1~2개월 앞서 조기 발생함이 확인되었다. 이러한 대발생은 약 4개월간 지속되었으며, 이는 해빙이 완전히 사라진 하계 기간보다 길어 북극 고위도 해역에서 UIBs가 중요한 현상임을 보여준다(Fig. 8). 또한 해빙이 후퇴한 이후 6월부터는 표층 아래 엽록소 최대층(Subsurface Chlorophyll Maximum, SCM)이 형성되어 약 4개월 동안 안정적으로 유지되었으며, 이는 기존 모델이 가정한 단기간 표층 대발생과 달리 지속적인 생산성 유지 메커니즘을 시사한다. KAMS 1 관측점에서 수직 엽록소 분포를 반영하여 추정한 연간 일차 생산량은 6.85 g C m-2yr-1로, 위성 기반 추정치의 두 배 이상으로 나타났다(Table 1). 이는 위성 관측이 SCM 층의 생산성을 포착하지 못해 북극해의 실제 탄소생산량을 심각하게 과소평가하고 있음을 의미한다. 반면, GOBAF 및 AOBAF 등 기존 생지화학 모델은 대발생의 시작 시기와 최대치를 평균 6~10주 늦게 예측하였으며, 최대 엽록소 농도 역시 실제 값보다 10배 이상 낮게 모의 되었다(Park et al., 2025). 이러한 불일치는 모델이 해빙아래 빛 환경과 수직 엽록소 분포의 역학을 적절히 매개변수화하지 못한 한계에서 비롯된 것으로 분석된다. 따라서 북극해의 일차 생산성과 탄소순환을 정확히 이해하기 위해서는 SCM 층과 UIBs의 역할을 포함한 연속적 현장 관측 기반의 모델 개선이 필수적이다. 특히 장기 계류 시스템은 위성 및 모델의 불확실성을 보완하고, 북극 고위도 해역의 계절적 생산성과 생태계 역동성 예측력을 강화하는 데 핵심적인 자료를 제공한다.
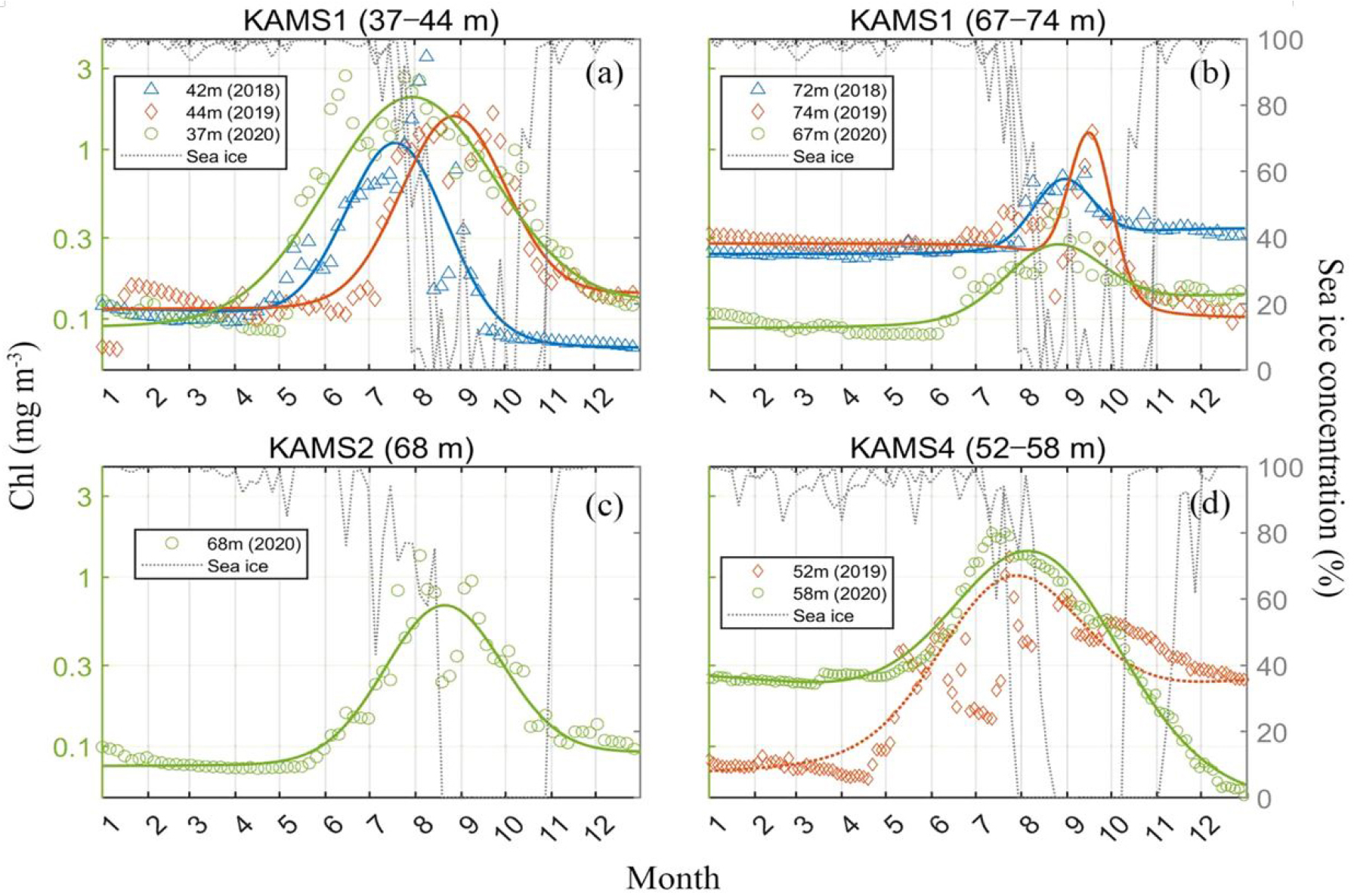
Fig. 8.
Seasonal cycle of chlorophyll-a concentrations (mg m−3, colored symbols) detected at individual Korea Arctic Mooring System (KAMS) stations by year. (a) The upper part of KAMS1 (37-44 m), (b) The lower part of KAMS1 (67-74 m), (c) KAMS2 (68 m), (d) KAMS4 (52-58 m). The gray dotted lines represent the daily time series of sea ice concentrations (%) around each mooring station. The colored lines represent the fitting lines for chlorophyll-a concentration estimated using the adjusted Gaussian curve fitting method (Figure from Ko et al., 2024).
Table 1.
Average primary production (PP) in August 2022, estimated using the Arctic-tuned vertically generalized productivity model (Cota et al., 2004). PP1 and PP2 are based on the vertical distribution of year-round chlorophyll-a (Chl-a) concentration from the Korea Arctic Mooring System 1 (KAMS1), with photosynthetically available radiation (PAR) derived from the Regional Arctic System Model (RASM), and satellite-derived PAR, respectively. PP3 is calculated using satellite-derived Chl-a concentration and PAR (Table from Park et al., 2025)
4.4 북극 대구(Boreogadus saida)의 분포 연구: eDNA 기법을 활용한 새로운 접근
북극 대구는 북극해 생태계 먹이사슬의 핵심 어종으로, 해빙 감소와 수온 상승 등 최근의 북극 온난화에 민감하게 반응하는 종이다. 북극해에서 북극 대구(Polar cod, Boreogadus saida)의 수평 및 수직 분포를 규명하기 위해 전통적 어류 채집 방법(링네트, 트롤, 핸드네트)과 eDNA 메타바코딩 기법을 병행한 연구가 최근에 보고되었다(Gwak et al., 2025). 전통적인 어류 채집 결과, 척치해 남부 및 동시베리아해에서 7~8월 초에 수집된 개체의 대부분은 연령이 0인 유어였으며, 8월 중순 이후에는 1~2살 연령의 개체가 주로 확인되었다(Fig. 9). 이러한 분포는 수온 및 먹이 이용 가능성과 밀접히 관련되며, 저온(-1.8~4°C) 수역에서 높은 밀도를 보였다. eDNA 메타바코딩을 통해서 베링해, 척치해, 동시베리아해역의 표층부터 최대 1804 m 깊이까지 북극 대구 DNA가 검출되었으며, 총 59개 정점 중 25개에서 양성 반응이 나타났다(Gwak et al., 2025). 이는 eDNA가 수평적‧수직적 분포를 동시에 파악할 수 있는 효과적인 수단임을 보여준다. 또한, 베링해에서는 eDNA가 검출되지 않아 이 지역에서의 출현이 제한적임을 시사하였다. eDNA는 전통적 조사 방법의 한계를 보완하면서, 고감도의 생물다양성 모니터링 도구로서 북극 생태계 연구에 유용함을 입증하였다. 북극 대구의 공간적 분포 및 서식 특성은 해양 온난화에 따른 북극 생태계 구조 변화의 지표로서 중요한 의미를 가지며, 향후 eDNA 기반 장기 모니터링 체계를 구축한다면 북극 어류군집의 변화 감시 및 생태계 예측에 핵심적인 정보를 제공할 수 있을 것이다.
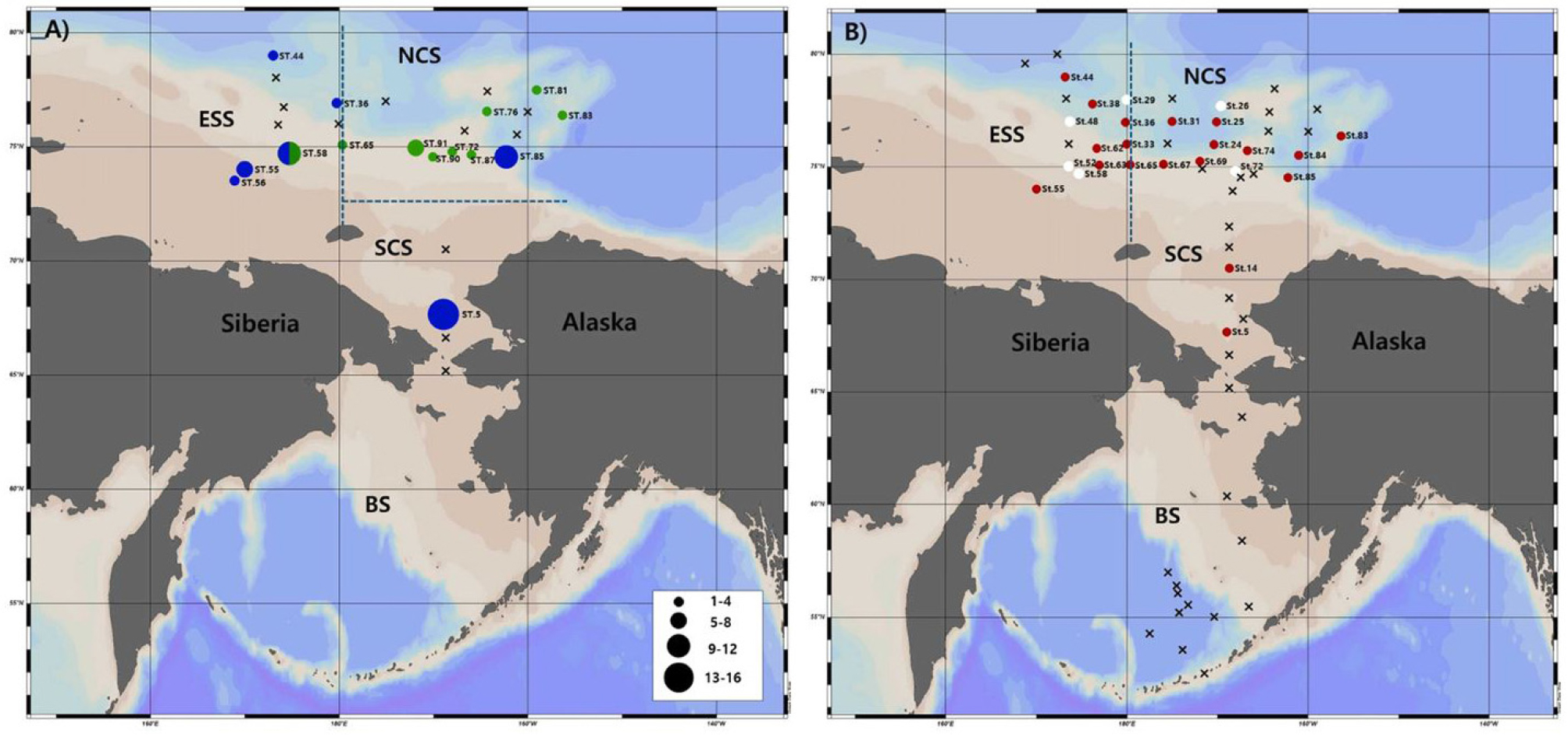
Fig. 9.
A map illustrating the locations where Boreogadus saida was collected using different nets (A), together with a map indicating the horizontal and vertical detection of B. saida eDNA (B). Horizontal differences in the number of B. saida are represented by the size of the circles (A). Blue indicates the location where B. saida was collected with ring net and/or frame trawl net, and green indicates the locations where it was collected with hand net (A). The red circles show where B. saida eDNA was detected just on the surface, whereas the white circles indicate locations where it was detected on both the surface and the bottom (B). BS: Bering Sea; SCS: southern Chukchi Sea, NCS: nothernChukchi Sea, ESS: East Siberian Sea. Black dashed lines are for separating SCS, NCS, and ESS. Reproduced from Gwak et al.(2025), Regional Studies in Marine Science, with permission from Elsevier.
4.5 서북극해에서 대서양 기원수 확장에 따른 생지화학 변화 현상 최초 규명
국제 다국적 공동탐사인 Synoptic Arctic Survey (SAS) 연구팀에서 관측된 서북극해의 생지화학적 이상 현상을 규명한 결과를 세계 최초로 제시하였다(Nishino et al., 2023). 연구진은 한국, 일본, 캐나다와 동시에 관측한 자료를 통합 분석하여, 척치고원 해역에서 전례 없는 용존산소가 낮고, 산성도가 높은 수괴를 발견하였다(Fig. 10). 이 현상은 대서양 기원수가 기존보다 동쪽으로 확장되면서, 동시베리아해 북부의 저산소 수괴가 척치 고원으로 북상한 결과로 나타났다. 이는 2010년대 중반 이후 북극진동(Arctic Oscillation, AO)이 양(+)의 위상으로 전환되며, 보퍼트 와류(Beaufort Gyre)가 수축하고, 동부 북극의 사이클론성 순환이 강화된 해양순환 구조 변화와 밀접히 연관되어 있다. 이러한 순환 변화는 동시베리아해 대륙붕 경사면의 저산소 수괴를 북쪽으로 이동시켜, 척치고원에서 용존산소 포화도가 50% 이하로 떨어지는 수괴를 형성하였다.
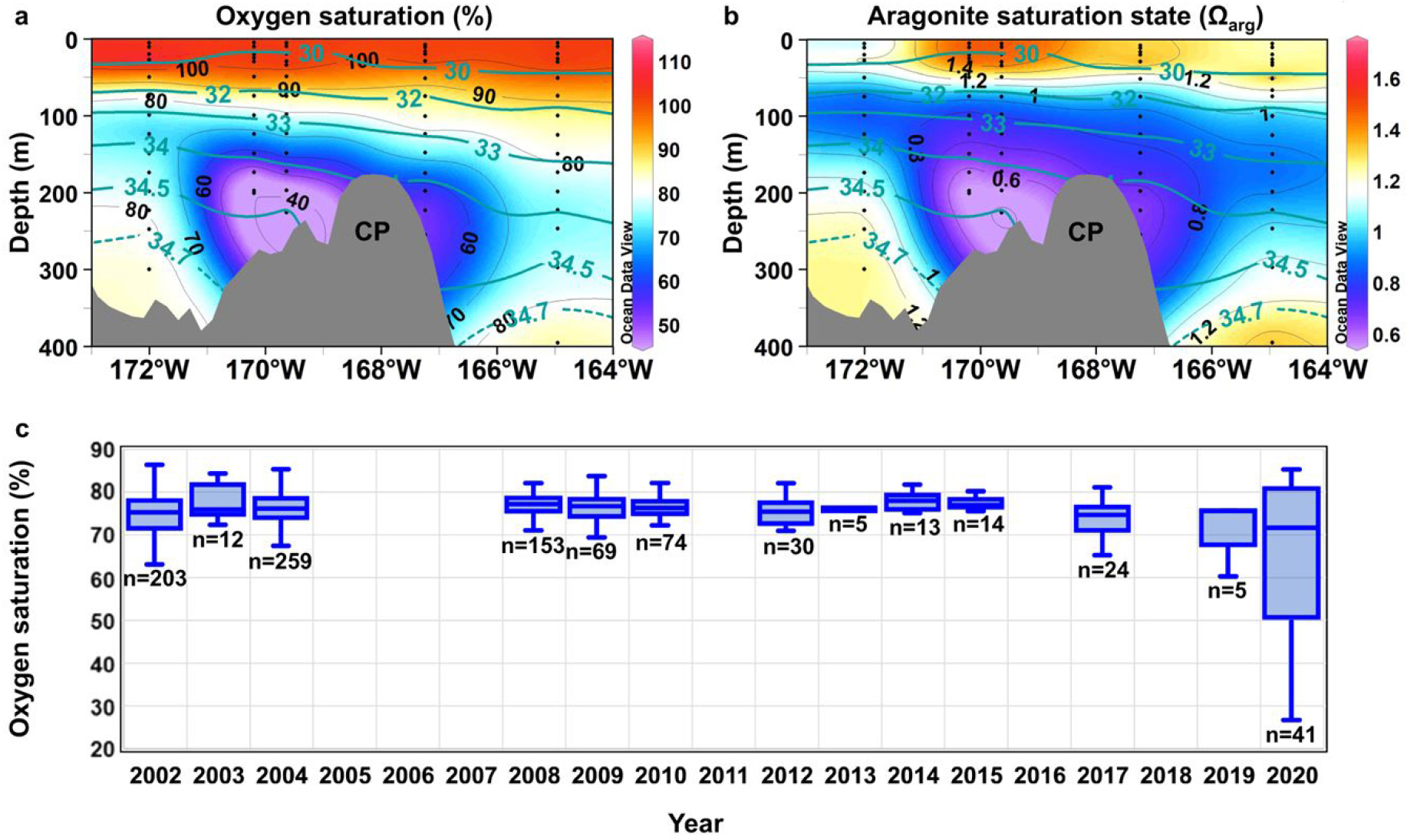
Fig. 10.
Water on the Chukchi Plateau in 2020 is characterized by anomalously low oxygen saturation and is corrosive to aragonite. a, b Vertical sections of oxygen saturation and aragonite saturation state, Ωarg, respectively, along 75° N across the Chukchi Plateau (CP; ~168° W) measured during the Research Vessel Miraicruise in 2020. Salinity contours are overlain, and black dots indicate the data points comprising each vertical section. c Box and whisker plot showing yearly variations in oxygen saturation. Boxes indicate the lower and upper quartiles. The horizontal line in each box represents the median oxygen saturation. Vertical lines extending from each box represent the minimum and maximum values recorded for that year. The number of samples for each year is shown below the minimum line (Figure from Nishino et al., 2023).
수치 모델 결과 이러한 저산소를 포함한 해수의 이동이 2017년 이후 지속적으로 발생했음을 발견하였으며, 해빙 감소와 대기순환의 변화가 대서양 기원수의 침투와 해양 생지화학적 이상 현상을 가속화했음을 보여준다(Nishino et al., 2023). 이러한 저산소‧산성화 수괴의 확장은 척치해 인근의 잠재적 어장 생태계에 부정적 영향을 미칠 수 있으며, 향후 중앙북극해 어업협정의 생태계 기반 관리 체계 수립 시 중요한 지표가 될 것으로 제시되었다. 따라서 본 연구는 서북극해가 대서양화의 영향권으로 빠르게 바뀌고 있음을 보여주는 결정적 증거로서, 북극 해양 순환 및 생지화학 시스템의 급격한 변화가 저산소‧산성화 위험 증가 및 생태계 구조 변동으로 이어질 수 있음을 시사한다.
4.6 서북극해에서 급속한 해양 산성화와 생지화학적 조절 요인
서북극해는 전 지구 해양 중 가장 빠른 속도로 해양 산성화가 진행되고 있는 지역으로, 이는 기후변화에 따른 해빙 감소, 담수 유입 증가, 그리고 수괴 교환의 변화와 밀접히 관련되어 있다(Qi et al., 2017; Qi et al., 2022; Kim et al., 2025). 최근 서북극해에서 산성화의 원인과 공간적 변동성을 정량적으로 규명함으로써, 북극 탄소순환과 생태계 변화를 이해하는 데 중요한 과학적 근거를 제시하였다(Kim et al., 2021b; Mo et al., 2022; Kim et al., 2025). 척치해와 동시베리아해 표층의 아라고나이트 포화도(Ωarag)는 평균 1.1~1.2 수준으로, 이미 포화 경계(Ω=1)에 근접하거나 일부 수역에서는 이보다 낮게 나타나고 있었다(Fig. 11). 이는 전 지구 해양 평균보다 빠른 속도로 산성화가 진행되고 있음을 의미한다. 특히 2016~2019년 사이, 부식성이 강한 태평양 기원수(Corrosive Pacific Water)의 기여도가 가장 높은 수심 50~300 m 구간에서 아라고나이트 불포화층(ΩAr < 1)이 급속히 확장되었으며, 이는 보퍼트 와류(Beaufort Gyre)의 담수 저장량 변화와 해빙 감소에 의해 조절되는 것으로 나타났다(Mo et al., 2022). 한편, 북극진동의 위상 변화 역시 산성화 강도를 결정하는 주요 요인으로 작용하였다. AO가 음(-)의 위상을 보일 때, 해수의 염분이 낮아지고 대기 CO2교환이 활발해져 산성화가 심화되며, 반대로 양(+) 위상에서는 대서양 기원수 유입이 강화되어 일시적 완화 경향을 보였다(Kim et al., 2025). 이러한 결과는 북극해의 산성화가 단순히 대기 중 CO2농도 증가에 따른 화학적 현상이 아니라, 해빙-담수유입-탄소순환-생물 생산성 간의 복합적 상호작용에 의해 결정되는 과정임을 보여준다. 특히 심층 불포화층의 확장은 탄산염 구조를 형성하는 패류나 저서생물(calcifier)들의 서식 환경을 위협하고, 결과적으로 북극의 탄소펌프 효율과 먹이망 구조의 변화를 유발할 수 있다(Mo et al., 2022). 따라서 본 연구 결과들은 북극 탄소 순환 및 산성화 예측 모델의 개선뿐 아니라, CAOFA 등 국제 해양 관리 체계에서 해양 산성화 지표를 포함한 장기 모니터링 체계 구축의 필요성을 강조하는 과학적 근거를 제공한다.
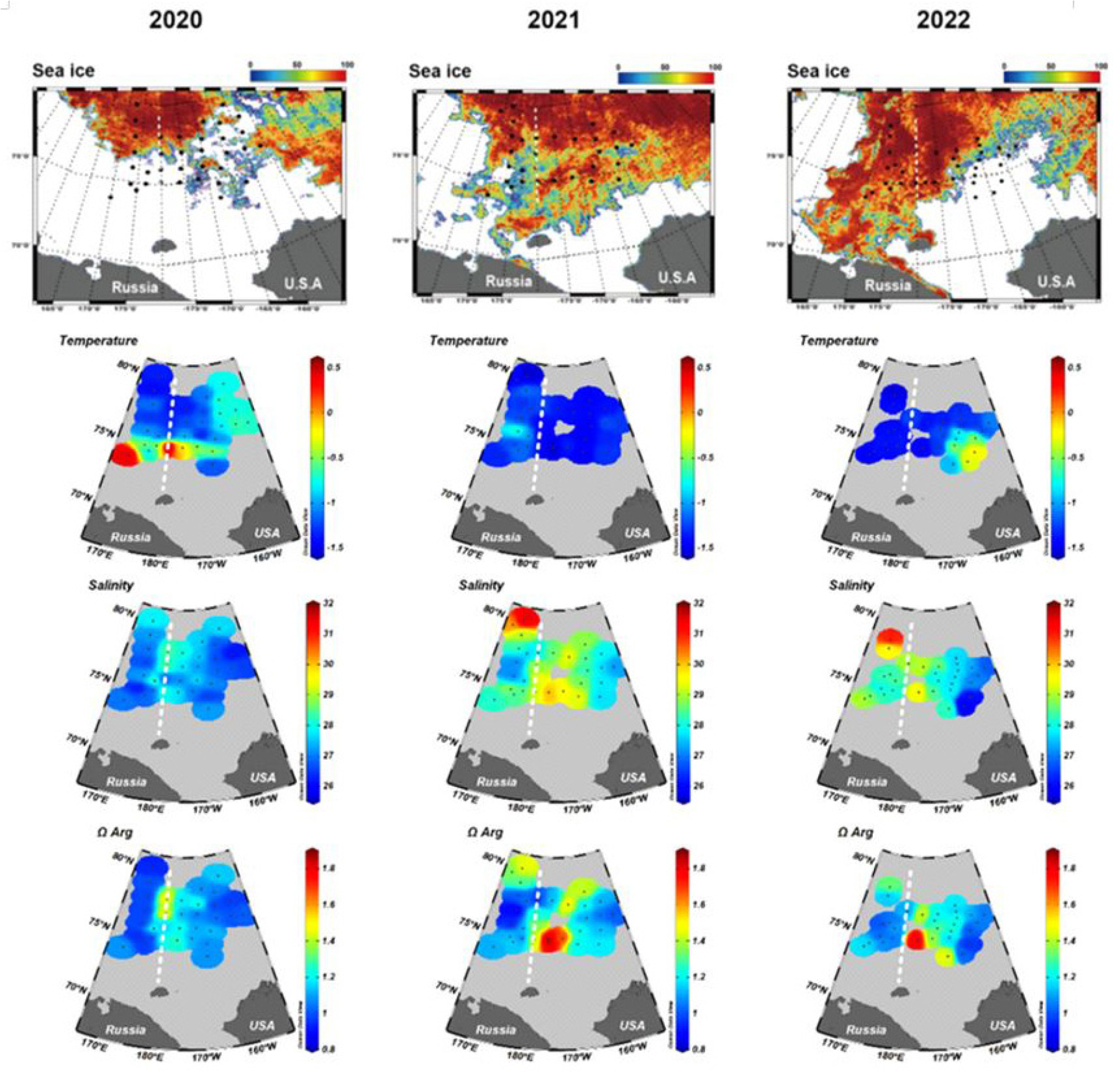
Fig. 11.
Surface distribution of sea ice, temperature, salinity and aragonite saturation state (Ωarag) from 2020 to 2022. White dotted lines indicate the boundary between the Chukchi marginal area and the East Siberia marginal area. Reproduced from Kim et al.(2025), Ocean Science Journal, with permission from Springer Nature.
4.7 북극 담수 유입 증가에 따른 생태계 반응 모델
급격한 북극 온난화로 인해 고위도 지역의 수문순환이 가속화되면서 북극해로 유입되는 강물 담수량이 크게 증가하고 있다. 이러한 담수 유입이 북극 해양 생태계, 특히 식물플랑크톤의 계절적 생산성과 분포 변화에 미치는 영향을 기후모델과 수치실험을 통해 분석하였다(Park et al., 2023). 현재 기후 조건에서 담수 유입이 증가하면, 봄철에는 해수의 염분이 낮아지고 결빙이 촉진되어 해빙 농도가 증가함으로써 광 이용 가능성이 감소하고 식물플랑크톤 생체량이 감소하였다. 반면 여름철에는 해빙 융해수 증가로 인한 혼합층 발달과 영양염 공급이 증가하면서 식물플랑크톤의 성장이 활성화되는 상반된 계절 효과가 나타났다. 이러한 결과는 담수 유입이 단순히 해양을 희석하는 작용에 그치지 않고, 해빙-빛-영양염 순환 간 복합적 상호작용을 통해 생산성을 조절함을 보여준다(Park et al., 2023).
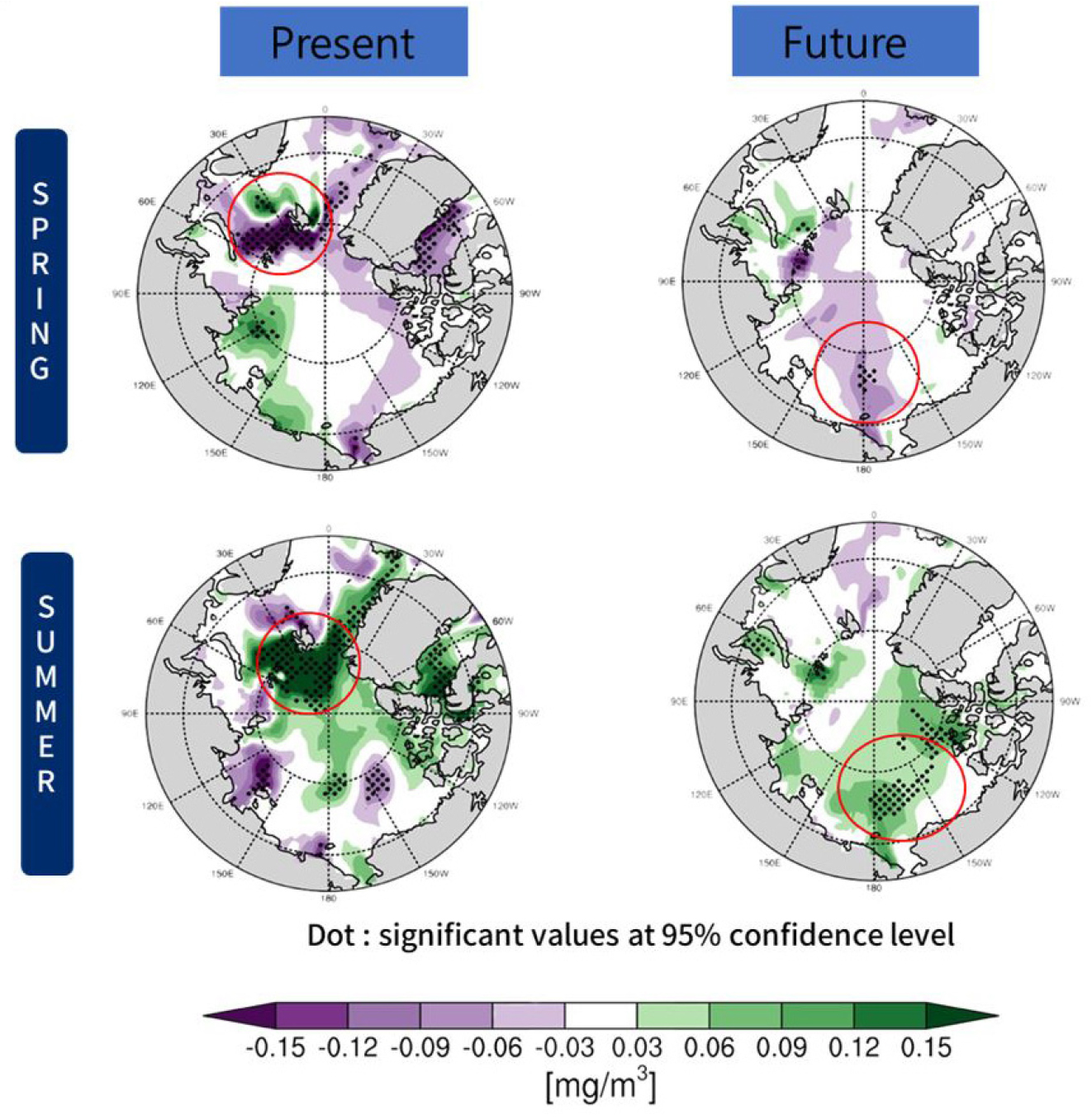
미래 기후 시뮬레이션에서도 유사한 계절적 반응이 지속되었으며, 특히 식물플랑크톤의 주요 반응 해역이 유라시아 분지에서 캐나다 분지와 동시베리아해로 이동하는 것으로 나타났다(Fig. 12). 이는 해빙 한계역(marginal ice zone)의 북상과 더불어, 북극 생태계의 중심이 되는 생산 해역이 동서로 변화될 가능성을 시사한다. 이러한 결과는 담수 유입이 북극 생태계 변동의 주요 조절인자임을 보여주며, 향후 어류자원 분포 변화 및 탄소순환 평가에도 중요한 영향을 미칠 것으로 예상된다. 따라서 본 연구는 강수 및 해빙 융해수 증가가 해양 생물생산성과 기후 되먹임을 동시에 조절하는 핵심 인자임을 규명하였으며, 이에 따라 해양생태-기후 상호작용을 정밀하게 모사할 수 있는 통합 모델의 개발 필요성을 강조하였다.
4.8 퇴적물 트랩을 활용한 서북극해 탄소플럭스 변동 연구
서북극해는 최근 해빙의 급격한 감소와 함께 일차 생산력, 생물학적 탄소펌프, 그리고 유기물 침강 과정의 변동이 두드러진 지역이다. 이러한 해양 생지화학적 변화를 이해하기 위해, 본 과제에서는 KAMS에 퇴적물 트랩(sediment trap)을 설치하여 계절별 입자물질 플럭스 변화를 관측하였다(Kim et al., 2021c; Gal et al., 2022). 본 연구에서는 해빙 하부에서의 해빙하 대번성(UIBs)과 그에 따른 유기탄소 및 생물기원 입자의 침강 특성을 파악하고, 해빙-해양-생태계 간의 상호작용을 정량적으로 규명하였다. 2017년 8월부터 2018년 8월까지 수행된 관측 결과, 동시베리아해 사면에서는 여름철(2018년 6~8월)에 입자 플럭스가 10배 이상 급증하였으며, 이는 풍부한 영양염을 함유한 대륙붕 기원의 냉수 유입에 의해 해빙 아래에서 UIBs가 발생된 결과로 나타났다(Fig. 13). 이 시기의 입자성 유기탄소(POC) 플럭스는 83 mgC m-2day-1에 달했으며, C:N 비율 상승과 규질조류의 증가가 동반되어 해빙조류 기원의 투명다당질(TEP) 생산이 중요한 역할을 했음을 보여준다. 반면 척치해 사면에서는 Anadyr 수의 안정적 유입으로 계절적 변동은 있으나 연간 변동이 적었고, 여름철 탄소 플럭스가 연간 총 유출의 약 75%를 차지하였다(Fig. 14).
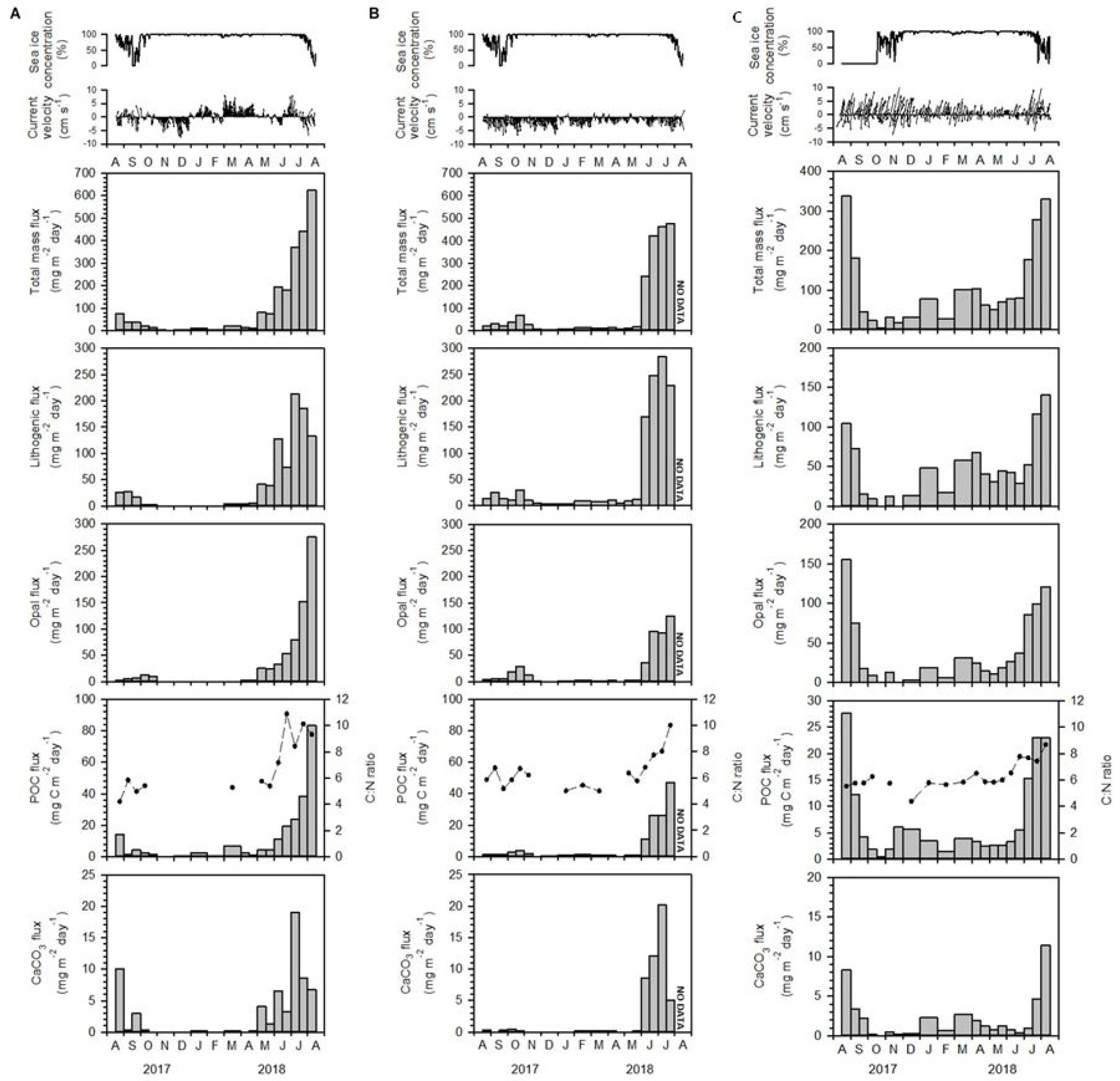
Fig. 13.
Sea-ice concentration, current velocity, total mass flux, POC flux (vertical bar), C:N ratio (dashed line), biogenic opal flux, CaCO3 flux, and lithogenic flux at water depths of 115 m (A) and 335 m (B) on the East Siberian Sea slope (KAMS 1), and 325 m in the Chukchi Sea (KAMS 2) (C) from August 2017 to August 2018 (This figure is modified from Kim et al., 2021c).
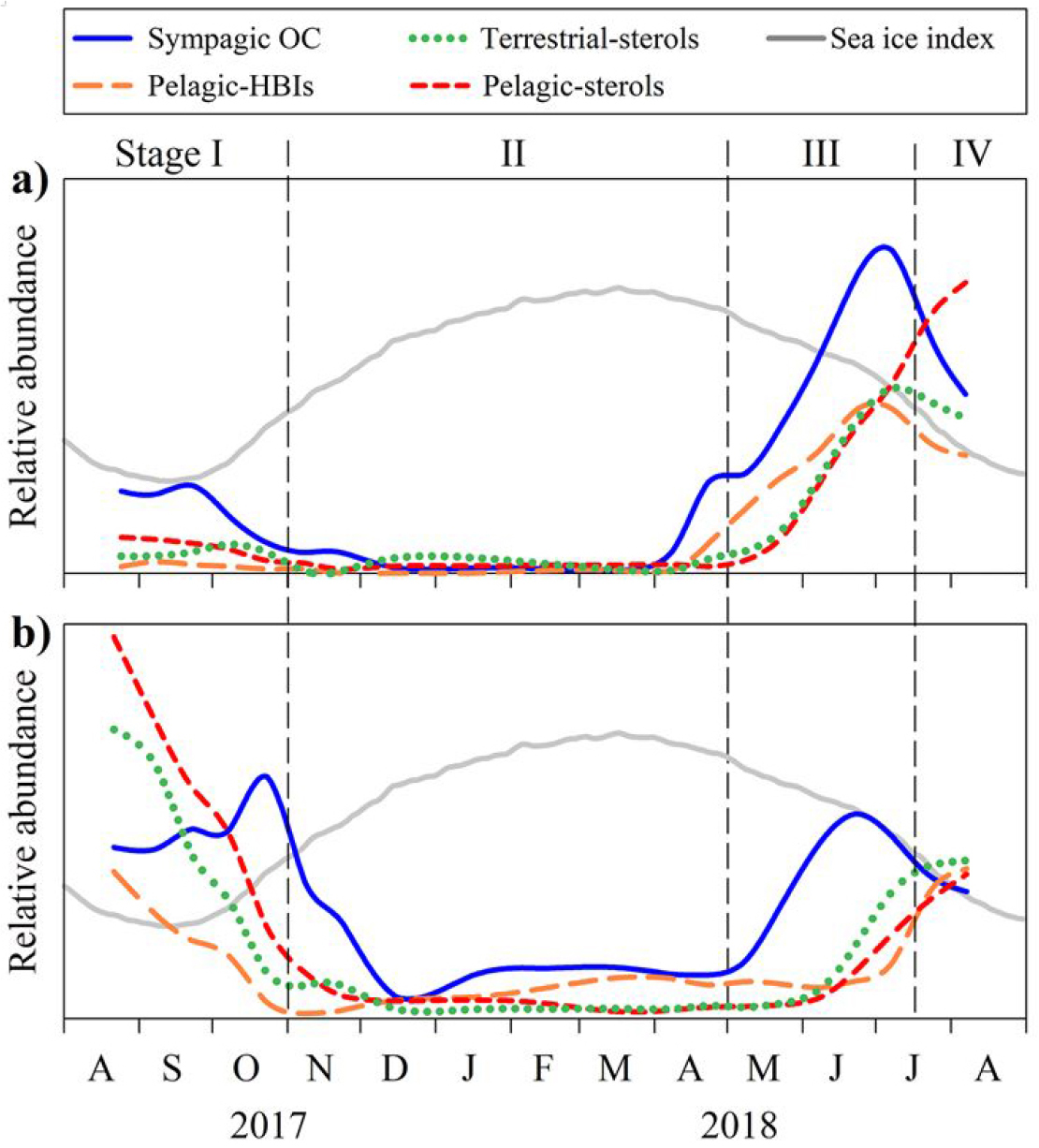
Fig. 14.
Three-point running average sympagicorganic carbon (OC) (solid blue line), pelagic highly branched isoprenoid (HBI) (long dashed orange line), terrestrial sterol (short dashed green line), pelagic sterol (medium dashed red line), and sea-ice index (obtained from the Northern Hemisphere from National Snow and Ice Data Center [NSIDC, https://nsidc.org]) values at the (a) KAMS1 and (b) KAMS2 stations. Reproduced from Gal et al.(2022), Journal of Geophysical Research: Ocean, with permission from AGU.
퇴적물 트랩에 포함된 지질 바이오마커(IP25, HBI, sterol 등) 분석결과, 해빙이 있는 시기에는 유기물 중 약 1~7%가 해빙 조류(sympagic algae)에서 기원한 것으로 추정되었다. 특히 동시베리아해역에서 UIBs 현상이 두드러지게 관찰되었고, 해빙이 존재하는 시기동안 유기탄소 플럭스는 척치해보다 약 5배 높았으며, 해빙하부에서 활발한 해빙조류 생산은 동시베리아해로 유입된 영양염 공급에 원인이 있는 것으로 나타났다(Fig. 14). 따라서 해빙이 녹지 않은 시기에 해빙하부 생태계의 생산성이 서북극해 탄소순환에서 점점 더 중요한 역할을 하고 있음을 시사한다. 이러한 결과는 퇴적물 트랩이 위성관측이 불가능한 겨울철 생산 및 탄소 이동을 규명할 수 있는 유일한 수단임을 보여준다. 본 연구는 급격히 변화하는 북극해 환경에서 해빙으로 덮혀 있는 시기에 해빙 하부 생태계가 탄소순환과 기후변화 완화에 미치는 잠재적 역할을 규명하는 데 중요함을 시사한다(Fig. 15).
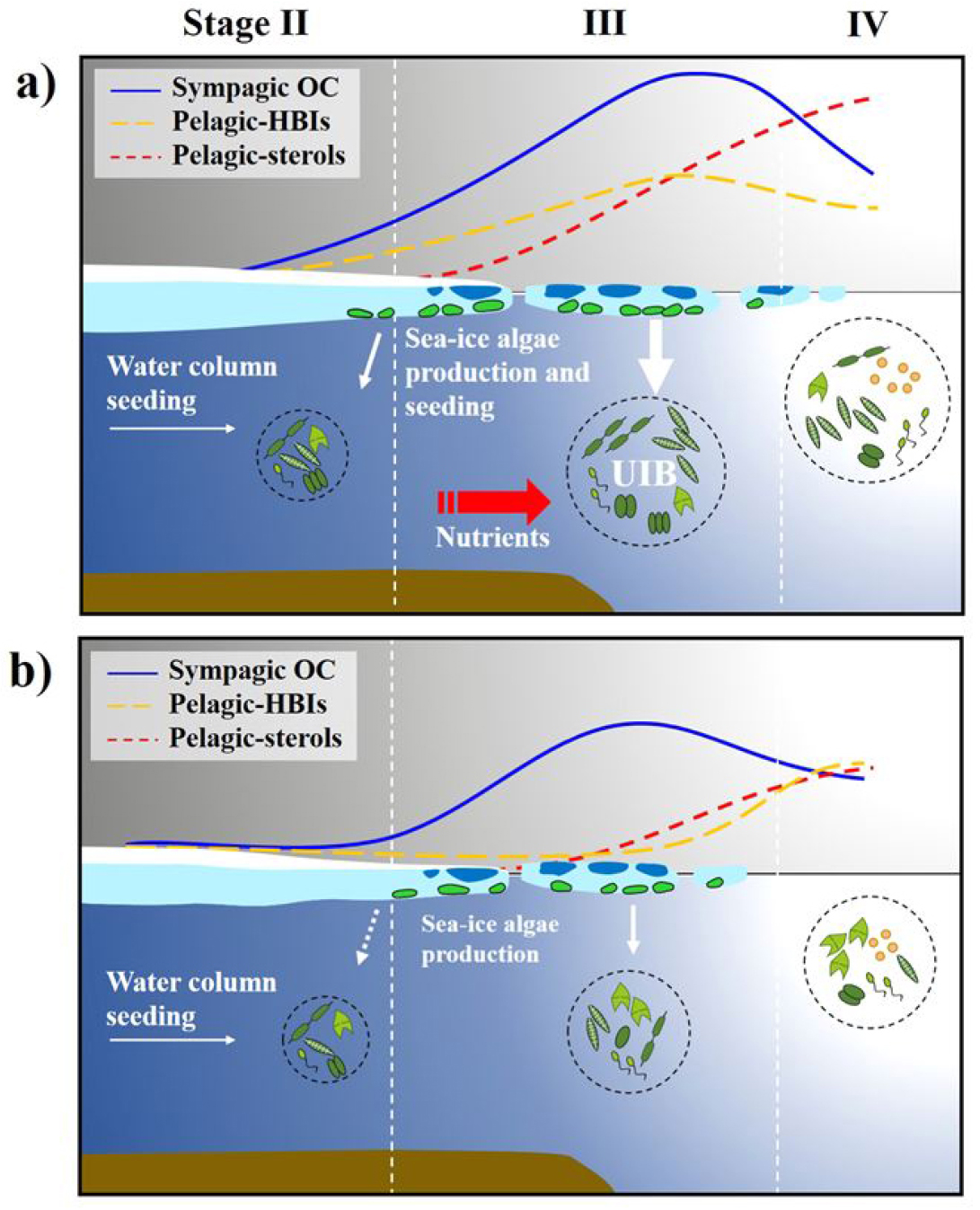
5. 결론 및 향후 전망
현재 북극해는 급격한 온난화로 인해 해빙이 빠르게 사라지고 있으며, 이는 기후 시스템과 해양환경, 생태계 전반에 중요한 영향을 미치고 있다. 급격한 해빙의 감소는 단순한 바다얼음이 사라지는 것이 아니라 기후‧환경 변화, 해양 생태계 구조변화, 수산자원 변동, 자원 개발, 북극항로, 그리고 국제 지정학적 관계까지 직결된 복합적 이슈이다. 이미 북극 해빙의 조기 소멸이 불가피한 현실로 다가오고 있는 만큼, 이에 대한 과학적 이해와 선제적 대응이 절실히 요구된다. 본 과제는 이러한 배경에서 서북극해를 중심으로 가속화되는 북극 온난화가 해양 환경과 생태계에 미치는 복합적 영향을 규명하고, 미래 변화를 예측할 과학적 기반을 마련하고자 하였다. 연구 결과, 북극해는 대서양화, 산성화, 담수화, 생태계 구조 변화가 동시에 진행되는 복합 전환기에 진입하였으며, 이는 북극 해양 시스템의 근본적인 변화를 의미한다. 이러한 변화는 전 지구 탄소순환과 기후 시스템의 불안정성을 증폭시키며, 북극이 더 이상 기후변화의 결과가 아닌 변화의 기점으로 작용하고 있음을 보여준다.
그러나 본 과제는 특정 해역에 집중된 조사로 인해 서북극해 전체를 대표하기 어렵다는 분명한 한계가 있으며, 서북극해의 특성만으로 북극해 전역의 변동성을 일반화하기에도 제한이 있다. 또한 조사 시기가 주로 하계에 국한되어 있어 일부 결과는 계절적 편향이 존재하며, 서북극해의 연중 변화 특성을 충분히 설명하는 데 어려움이 있다. 더불어 본 과제는 다양한 자료를 다루고 있으나, 결과들이 독립적으로 제시되어 해양환경 변화와 생태계 변동을 정량적으로 연결하는 통합적 생지화학‧생태 모델링이 미흡한 한계가 있다. 이러한 문제점을 보완하기 위해서는 급변하는 북극해 시스템을 연중 안정적으로 관측할 수 있는 장기 모니터링 체계의 고도화와 더불어, 물리-화학-생태계를 아우르는 통합 예측 역량 강화가 필수적이다. 이를 위해 해빙-해양-생태계를 아우르는 다중센서 기반 장기 계류 시스템과 준실시간 관측망을 확대하고, 현장 기반 자료를 활용한 해양-생지화학-생태계 통합 모델의 정교화가 필요하다. 특히 담수 유입, 산성화, 해빙 감소가 복합적으로 작용하는 북극해의 생산성 변화 메커니즘을 규명하고, 이를 예측 가능한 시뮬레이션 체계로 발전시키는 것이 향후 핵심 과제가 될 것이다.
이와 함께 국제 공동연구와 데이터 상호운용을 강화하여, 한국이 북극해 연구의 국제 과학 허브로서 역할을 수행해야 한다. KAOS와 같은 데이터 플랫폼을 기반으로 한 개방형 정보 공유, CAOFA 등 국제 협정 이행을 지원하는 과학적 근거 구축은 지속 가능한 북극 관리와 국제 협력의 중심축이 될 것이다. 특히 2029년에 건조될 차세대 쇄빙연구선은 이러한 연구를 획기적으로 확장할 수 있는 핵심 인프라가 될 것이다. 북극항로에 대한 접근성 향상과 북극해 전역에서 통합 관측이 가능해지면, 이를 통해 한국은 향후 북극연구와 국제 기후변화 대응에서 핵심적이고 과학적 리더십을 강화할 수 있을 것이다. 국가 차원의 지속적인 투자와 연구 인프라 활용은 극지 과학기술 인력 양성 및 국가 과학 역량의 구조적 강화로 이어져, 향후 북극해 연구의 패러다임을 선도하는 동력이 될 것으로 전망된다.
