

1. 서 론
2. 재료 및 방법
2.1 퇴적물 및 해수 채집
2.2 수조 설계 및 제작
2.3 중형저서동물 군집 변동
2.4 먹이생물 배양: 박테리아 및 저서성 규조류
3. 결 과
3.1 Exp. 1 : 자연 상태 조건에서의 중형저서동물 군집 구조 변화
3.2 Exp. 2 : 먹이 공급 유무에 따른 중형저서동물 군집 구조 변화
3.3 Exp. 3 : 먹이유무 및 광조건에 따른 군집 구조 변화
4. 고 찰
1. 서 론
중형저서동물은 해양 퇴적물의 입자와 입자 사이에서 서식하는 간극동물로써, 망목 1 ㎜를 통과하여 32 ㎛ 체에 걸러지는 크기에 들어가는 생물이다. 해양 생물의 그룹 중에서는 문 준위의 가장 다양한 분류군이 포함되어 있으며, 단위면적당 서식밀도가 높게 나타난다(Giere, 2009). 중형저서동물은 어류, 패류 등 다양한 대형동물의 먹이원이며(De Morais and Bodiou, 1984; Castel, 1992; Ceccherelli et al., 1994), 유기물, 미생물, 원생생물들을 주로 먹이원으로 이용한다(Pergent et al., 1994; Duarte and Cebrian 1996; Gwyther 2003; Torres-Pratts and Schizas 2007; Mascart et al., 2013, Mascart et al., 2015). 이처럼 중형저서동물은 저서생태계 내 먹이사슬의 중간 연결고리 역할을 하는 중요한 위치를 차지하고 있다(Giere, 2009; Woodward, 2010; Zeppilli et al., 2015). 이러한 중형저서동물은 대형저서동물에 비해 세대교번이 짧고, 환경 변화에 매우 민감하게 반응하는 생태적 특성이 있다(Bongers and Ferris, 1999). 또한 중형저서동물은 퇴적물의 생물혼탁작용(bioturbation) 또는 미생물의 활동을 자극하여 유기물 분해를 촉진시키는 역할 등 주변 환경의 변화에도 일조하는 것으로 알려져 있다(Aller and Aller, 1992; Giere, 2009). 이러한 특성들로 인하여 중형저서동물은 해양 생태계의 변화를 감지하거나 예측하는데 매우 유용하게 활용되고 있다(Jessup et al., 2004; Nascimento et al., 2011, Nascimento et al., 2012; Bonaglia et al., 2014). 그러나 중형저서동물은 구성 분류군의 특성을 파악하기 위해서는 고배율 현미경을 통해서만 관찰이 가능하고(Balsamo et al., 2010), 이로 인해 이들 분류군에 대한 정보가 매우 제한적인 상태이다(Giere, 2009). 최근 연구에서 그 크기때문에 중형저서동물은 해양 저서생태계의 주된 대상생물인 대형저서동물의 생물다양성과 생태적인 기능에 영향을 미치는 것으로 보고되었다(Piot et al., 2014; Pusceddu et al., 2014b). 이와 같이 중형저서동물의 저서생태계 내에서 기능적인 면은 지금까지의 연구들에서 추정되어진 것 보다 더욱 복잡할 것으로 예상되며, 그렇기 때문에 향후 해양생태계 내에서 중형저서동물의 역할, 기능 등의 중요성에 대한 연구의 필요성은 매우 높다고 볼 수 있다(Balsamo et al., 2010).
이러한 중요성이 있음에도 불구하고, 중형저서동물은 그 크기가 매우 작고, 군집을 구성하는 각 분류군들 마다 생리·생태적 특성이 매우 복잡하고 다양하며, 수명도 일주일에서 일년 이상까지 분류군마다 다양하여 군집 규모의 배양 실험을 수행하기에는 어려운 특성을 가지고 있기 때문에 최근까지도 중형저서동물 군집 규모의 배양 연구는 거의 이루어지지 못하고 있는 실정이다(Higgins and Thiel, 1988). 중형저서동물이 실험실 내에서 자유롭게 배양이 가능한 시스템을 갖추게 된다면, 실험을 위하여 매번 현장에 가서 생물 시료를 채집해야하는 시간적·경제적 소모를 줄일 수 있다. 또한 이를 활용하여 다양한 목적의 실험에 활용함으로써 그 가치를 더욱 높일 수 있는 등 그 활용도는 매우 높아 질 것이며, 중형저서동물의 생리적·생태적 역할에 대하여 더욱 심도있는 연구를 수행 할 수 있다. 일반적으로 중형저서동물의 높은 다양성이나 서식밀도는 서식처의 온도나 염분도, 산소의 양, 퇴적물 입도 등 여러 환경 요인의 영향을 많이 받는 것으로 알려져 있다(Ingels et al., 2011; Lizhe et al., 2012; Ngo et al., 2013; Pusceddu et al., 2014a).
본 연구에서는 여러 환경 요인들 중에 중형저서동물 군집의 다양한 생물들에게 환경 조건으로 가장 중요한 요소들인 먹이의 종류와 광조건을 배양 조건으로 설정하였다. 조건들은 실험의 단계별로 호조건과 부적합 조건들을 새로 설정하고 배제하였고, 여러 조건을 설정한 실험을 통하여 실험실 내에서 중형저서동물 군집 규모의 배양을 가장 이상적으로 유지할 수 있는 조건을 찾아내고자 본 연구를 수행하였다.
2. 재료 및 방법
2.1 퇴적물 및 해수 채집
중형저서동물 군집 규모의 배양 실험에 사용할 퇴적물은 태안 만리포 조간대 지역(Fig. 1)에서 확보하였다. 퇴적물이 채집 된 지역은 중형저서동물 분류군이 주변 지역에 비하여 상대적으로 다양한 지역이다(Kang et al., 2016). 또한 퇴적상이 사질로 구성되어 있어 수조 배양에서 이용시 퇴적물의 부패가 니질의 퇴적물보다 느리게 진행되는 장점이 있다.
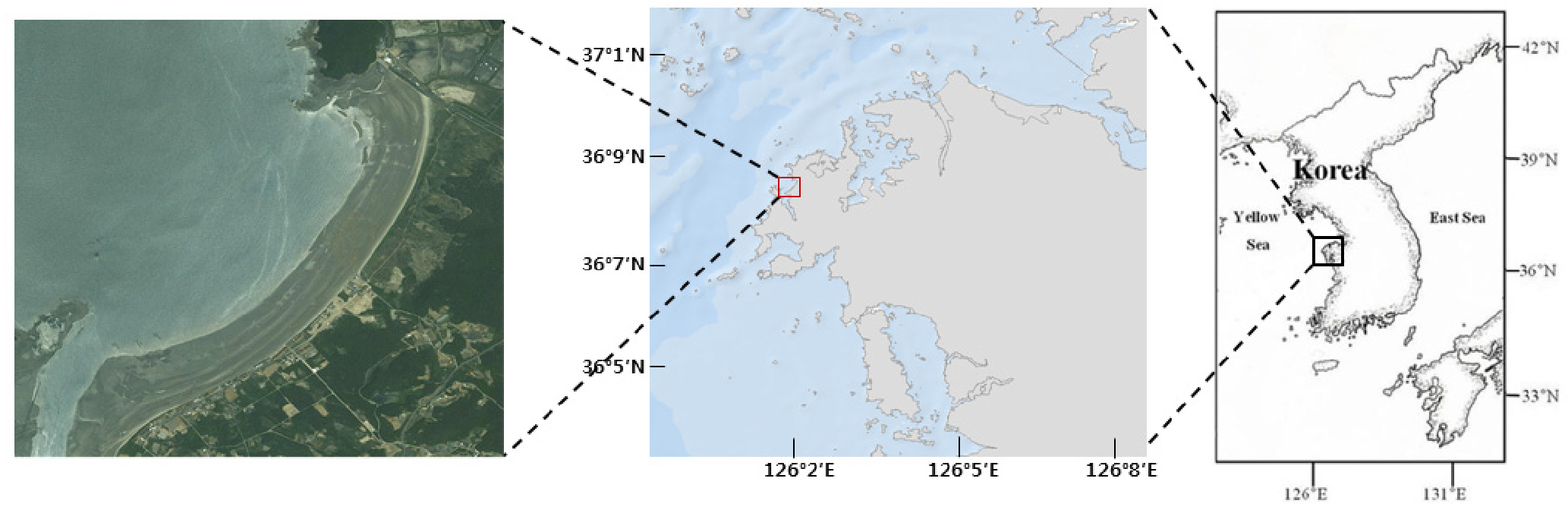
퇴적물 채집은 2013~2014년에 이뤄졌으며, 퇴적물은 일반적으로 중형저서동물이 가장 많이 서식하고 있는 표층에서 5 ㎝까지 채취하였다. 채집된 퇴적물을 실험실로 이동시에는 가능한 자연 상태의 환경을 유지하기 위하여, 저온(4~10℃)의 아이스박스에 넣고 산소 공급을 하면서 이동하였다. 또한 퇴적물이 채집된 장소에서 해수를 같이 채집하여 실험실로 운반한 뒤, GF/F (GFF-047, Whatman®) 여과지로 해수를 여과시킨 후 배양에 사용하였다.
2.2 수조 설계 및 제작
중형저서동물 군집의 배양을 위한 수조는 직접 설계 제작하였으며, 수조의 크기는 600×450×450 (W×D×H/㎜)로 총 4개를 제작하였다. 수조 내 하단에는 중형저서동물이 배양될 퇴적물의 부패를 방지하기 위해 해수만 통과되고 퇴적물은 통과되지 못하는 구조물을 설치하였다. 배양수조 내의 해수 순환은 수조 상층의 해수를 펌프로 빨아들여 여과시킨 후, 이 여과된 해수가 퇴적물 하부로 유입되어 상층 방향으로 공급되게 제작하였다. 이러한 구조의 수조를 이용하여 퇴적물 내의 해수 순환을 시켜줌으로써 퇴적물의 부패를 최대한 방지하였다(Fig. 2).
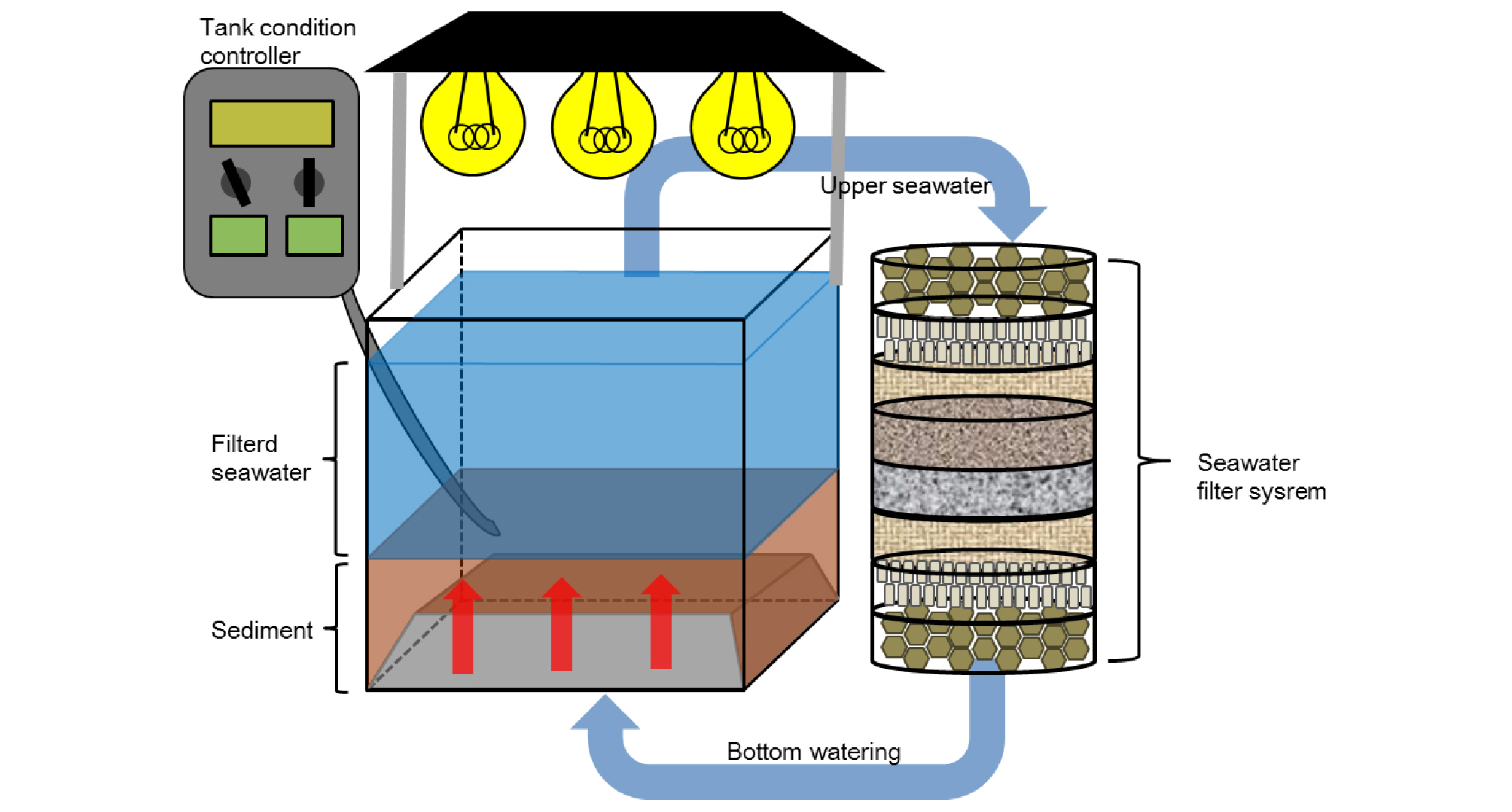
또한 배양 수조 내의 환경 조건은, 채집 지역인 만리포 해변의 자연 상태와 유사한 용존산소(7.0~8.0 ppm), 염분(29~31 psu), 온도(19~20℃), 광주기(12 hr.) 조건을 설정 하였다. 수조 내 염분 농도를 유지하기 위한 장치로, 해수가 증발이 되어 수층이 낮아지면 자동으로 증류수가 공급되는 장비를 설치하였다. 또한 각 수조에 각각의 온도와 광주기를 설정할 수 있는 조절장치를 장착하였으며, 각 수조의 온도와 용존산소를 확인할 수 있는 모니터링 장비를 설치하였다. 또한 수조 내에 다공성의 우드스톤이 장착된 유회수기(skimmer)를 설치하여 중형저서동물 군집이 배양되는 동안 해수 내 유기물 농도가 높아지지 않도록 유기물 농도를 조절하였다. 더불어 산소 농도 유지를 위해 수조 내에 산소를 공급해주는 기포기를 배치하였다. 4개의 배양 수조는 온도와 습도 조절이 가능한 컨트롤 챔버 내에 설치되어 실험을 진행하였다. 채집된 퇴적물은 자연 상태에서의 높이와 마찬가지로 수조의 바닥에서 약 5 ㎝ 높이가 되게 채워졌으며, 퇴적물과 함께 약 80 ℓ의 여과 해수로 수조를 채워 실험을 진행하였다.
2.3 중형저서동물 군집 변동
수조 내 중형저서동물의 서식밀도 변화를 파악하기 위한 시료의 채집은 각 수조에서 주사기 코어러를 이용하여 3 반복으로 일주일에서 한달 간격으로 수행되었다. 중형저서동물 군집 배양 실험은 총 세 번에 걸쳐 조건을 달리하여 진행하였다. 첫 번째 실험(Exp. 1)은 채집 지역과 같은 용존산소, 염분도, 온도를 유지하며 12 시간의 광주기를 주고 퇴적물에 어떠한 첨가도 없이 중형저서동물 군집의 변화를 살펴보았다.
두 번째 실험(Exp. 2)에서는 첫 번째 실험의 조건에 중형저서동물 군집의 먹이생물인 미동정 박테리아 5종과 분양받은 갈조류(Isochrysis galbana), 그리고 녹조류(Tetraselmis suecica)를 공급한 실험구(Exp. 2, 수조 B)와 공급하지 않은 대조구(Exp. 2, 수조 A)로 나누어 실험을 수행하였다.
마지막 실험(Exp. 3)은 용존산소, 염분도, 온도는 선행된 실험과 같지만 암배양과 명배양(12 시간 주기), 먹이(박테리아, 저서성규조류)의 공급 유무에 따른 중형저서동물 군집 구조 변동을 살펴보았다(Fig. 3, Table 1). 자연상태와 유사한 조건에서 빛조건을 제외한 암배양으로 실험한 수조 A, 자연상태와 유사한 조건에서 빛조건을 제외하고, 박테리아와 저서성 규조류를 먹이로 공급한 수조 B, 자연상태와 유사한 조건으로 실험한 수조 C, 그리고 자연 상태와 유사한 조건에서 박테리아와 저서성 규조류를 먹이로 공급한 수조 D로 조건을 설정하여 배양을 진행하였다. 수조 A와 C에는 만리포 해변에서 서식하는 박테리아와 저서성 규조류(diatoms)를 배양한 후, 혼합하여 중형저서동물 군집의 먹이로 공급하였으며 수조 B와 D에는 먹이를 공급하지 않는 조건을 설정하였다(Exp. 3, 수조 A~D).
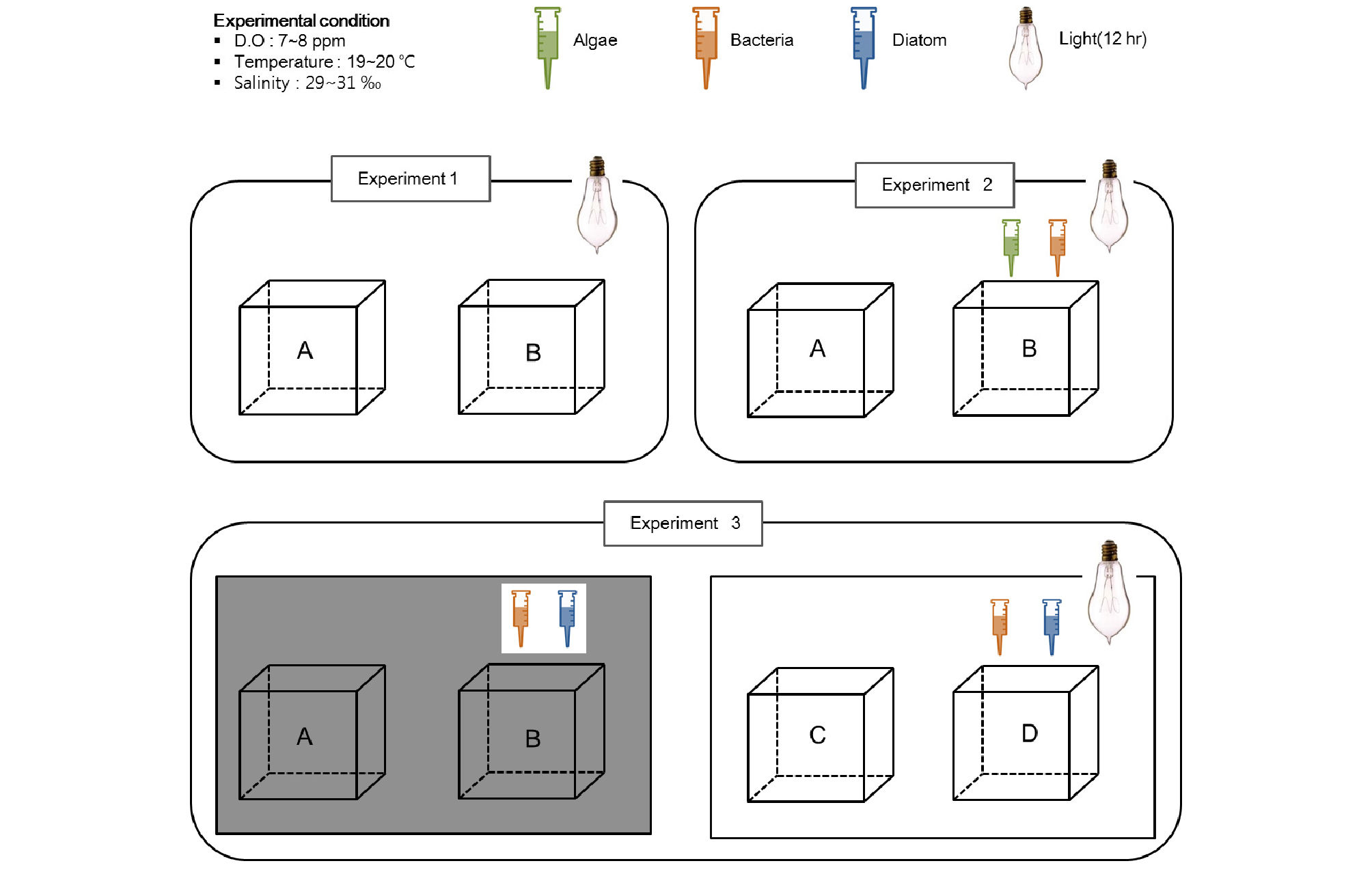
Fig. 3.
Overview of the three experimental conditions: Experiment 1) Changes under simulated natural conditions (A, B). Experiment 2) Changes according to no-food (A) or food containing algae and bacteria (B). Experiment 3) Changes according to no-light with no-food (A) or food containing bacteria and diatoms (B) and light with no-food (C) or food containing bacteria and diatoms (D).
Table 1. Culture conditions of meiofauna community
Margalef(1958)의 정보이론에서 도출된 Shannon-Wiener 함수에 의해 유도된 종다양지수(H’)를 사용하였다. 또한 각 실험의 조건에 따른 중형저서동물 군집의 생존시간에 대한 유의한 차이를 파악하기 위하여 SPSS 19를 이용하여 일원분산분석(One-way analysis of variance, One-way ANOVA)과 이원분산분석(Two-way analysis of variance, Two-way ANOVA)을 실시하여 유의성을 검정(95% 신뢰수준)하였다.
2.4 먹이생물 배양: 박테리아 및 저서성 규조류
중형저서동물의 먹이생물은 퇴적물이 채집된 지역과 같은 조건을 유지하기 위하여 만리포 지역의 박테리아와 규조류 채집하여 배양하였다. 박테리아의 배양은 채집해온 퇴적물과 해수를 혼합한 뒤, 퇴적물을 가라앉히고 상층의 해수를 분리해 1/10⁴로 희석하여 멸균해수로 제작한 1.5% 한천배지에 접종하고 유리구슬을 이용하여 도말한 뒤, 30℃에서 배양하였다. 배양 1주일 후, 박테리아의 단일 배양을 위하여 백금이(loop)를 이용하여 발생한 콜로니(colony)를 하나씩 채취해 멸균해수에 효모추출물(yeast extract)를 첨가한 1.5% 한천배지에 3분 도말한 후, 30℃ 배양기 내에서 배양하였다. 4일 후 단일 배양된 박테리아들을 콜로니의 형태와 색깔 등에 따라 분류한 뒤, 우점하는 5종을 선별하여 대량배양 하였다. 대량배양은 멸균해수에 2%의 효모추출물을 첨가해 제작한 액체 배지를 멸균된 일회용 유리바이알(BD Falcon™)에 약 4~5 ㎖씩 넣어 주고 박테리아를 접종하여 박테리아 배양기에서 3일 동안 배양하였다. 5종의 박테리아는 E.P tube에 1~2 ㎖ 씩 넣어 동량의 40% 글리세롤(glycerol)을 첨가하여 –70℃에서 동결 보관하였다.
저서성 규조류의 배양은 GF/F 여과지로 여과한 해수를 121℃에서 15분 동안 멸균시킨 뒤, 여과해수 1ℓ당 f/2 (GUILLARD’S, without silicate)를 200 ㎖의 비율로 혼합한 배양액을 제작하였다. 만리포 해변에서 채집해온 퇴적물에서 미동정 저서성 규조류를 분리해낸 뒤, 세포 배양 플라스크(CELLSTAR®, greiner bio-one) 또는 플라스틱 자(jar)에 배양액과 함께 넣어 혼합 배양하였다.
3. 결 과
3.1 Exp. 1 : 자연 상태 조건에서의 중형저서동물 군집 구조 변화
첫 번째 실험(Exp. 1)은 실험의 정밀도를 높이기 위해 두 개의 수조(A, B)에 동시 반복실험 하여 결과 값을 도출하여 비교하였다. 실험 결과, 수조 A와 수조 B의 서식밀도 변동은 상당히 비슷한 경향을 나타냈다(Fig. 4). 수조 A와 B 모두, 배양 약 7주 내에서는 수조 A에서 최고 68 개체/10 ㎝², 수조 B에서 79 개체/10 ㎝²로 일시적으로 급격히 증가하는 경향을 보였지만 그 이후 양쪽 수조 모두에서 서식밀도가 급격히 감소하는 모습을 보였다. 배양 12주가 지나서는 초기 서식밀도에 비하여 약 50%의 서식밀도로 감소하였으며, 최저 서식밀도는 수조 A에서 29 개체/10 ㎝², 수조 B에서 21 개체/10 ㎝²로 배양 종료 시점인 20주까지 비슷한 서식밀도로 유지되었다(Table 2).
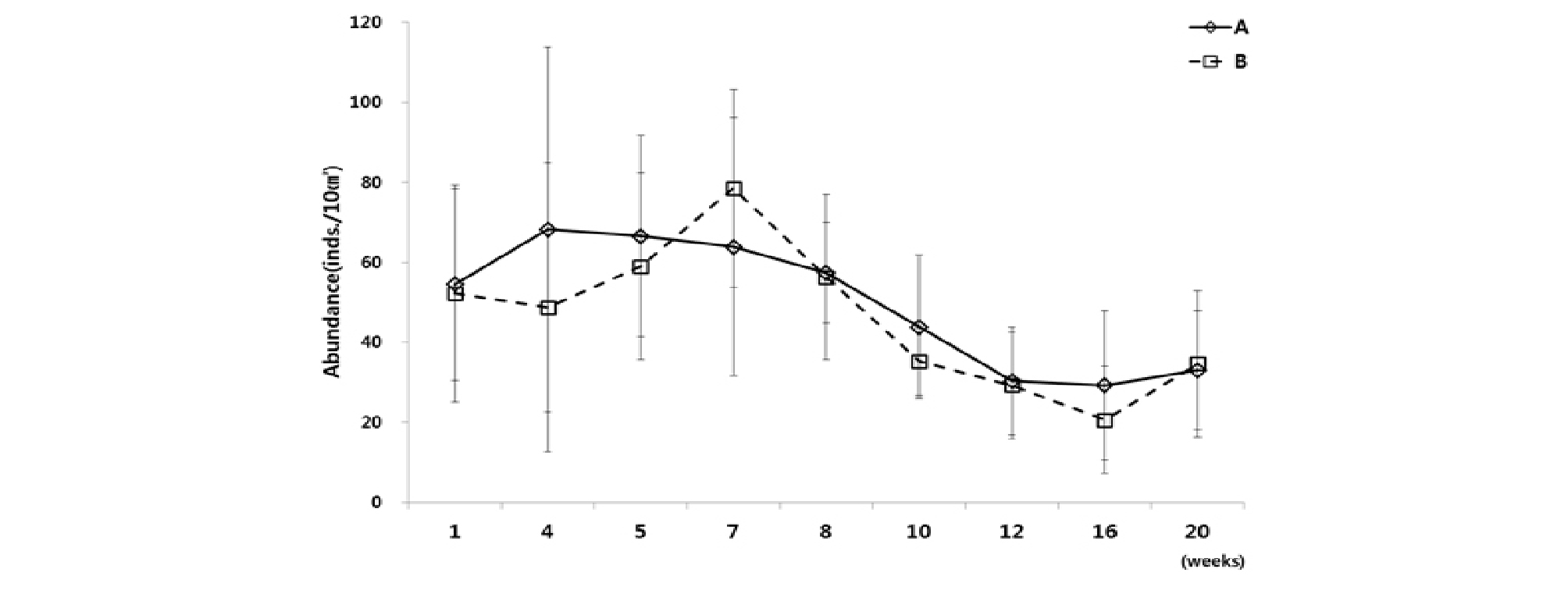
Table 2. Total density of meiofauna at experiment 1 (Light and no-food)
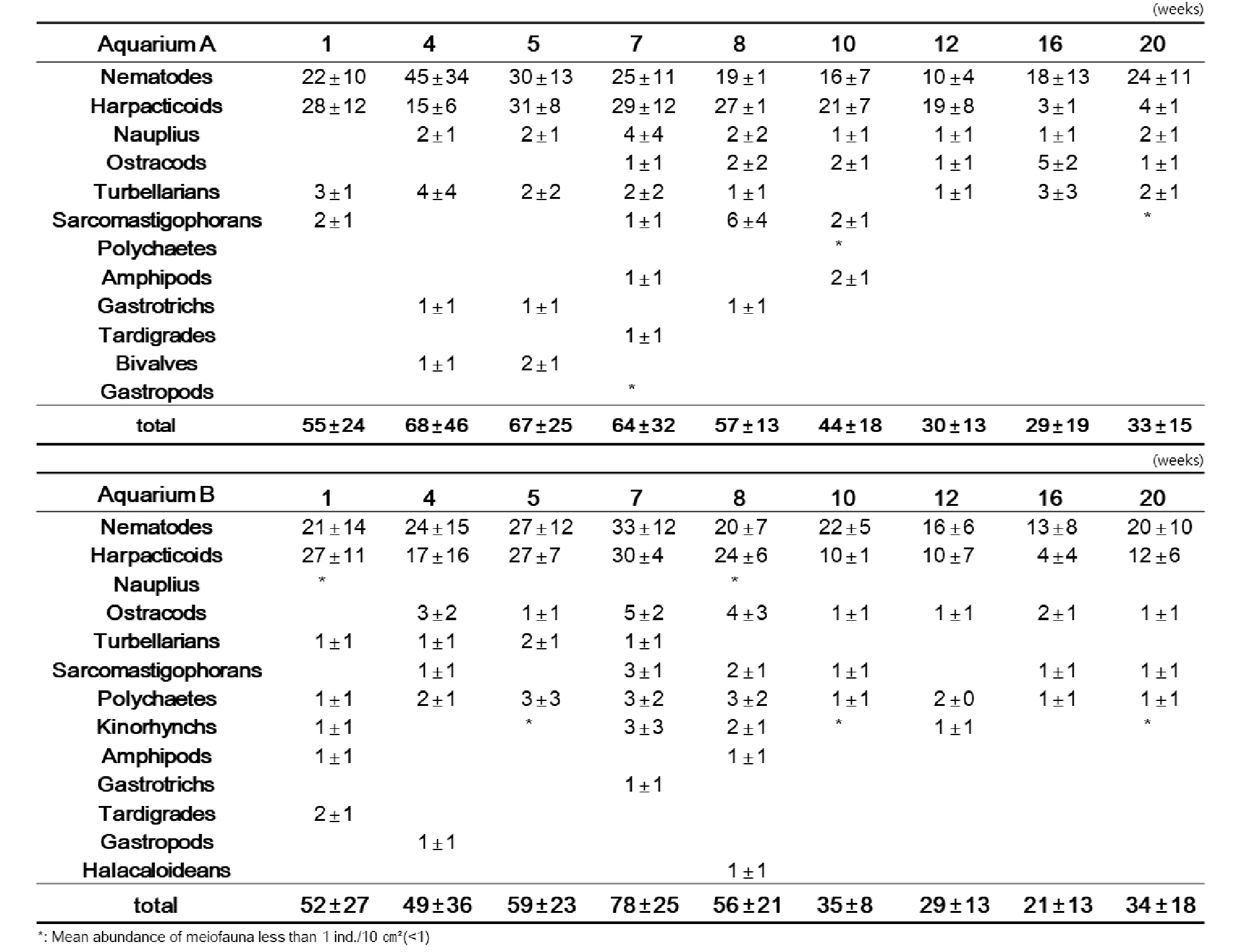 |
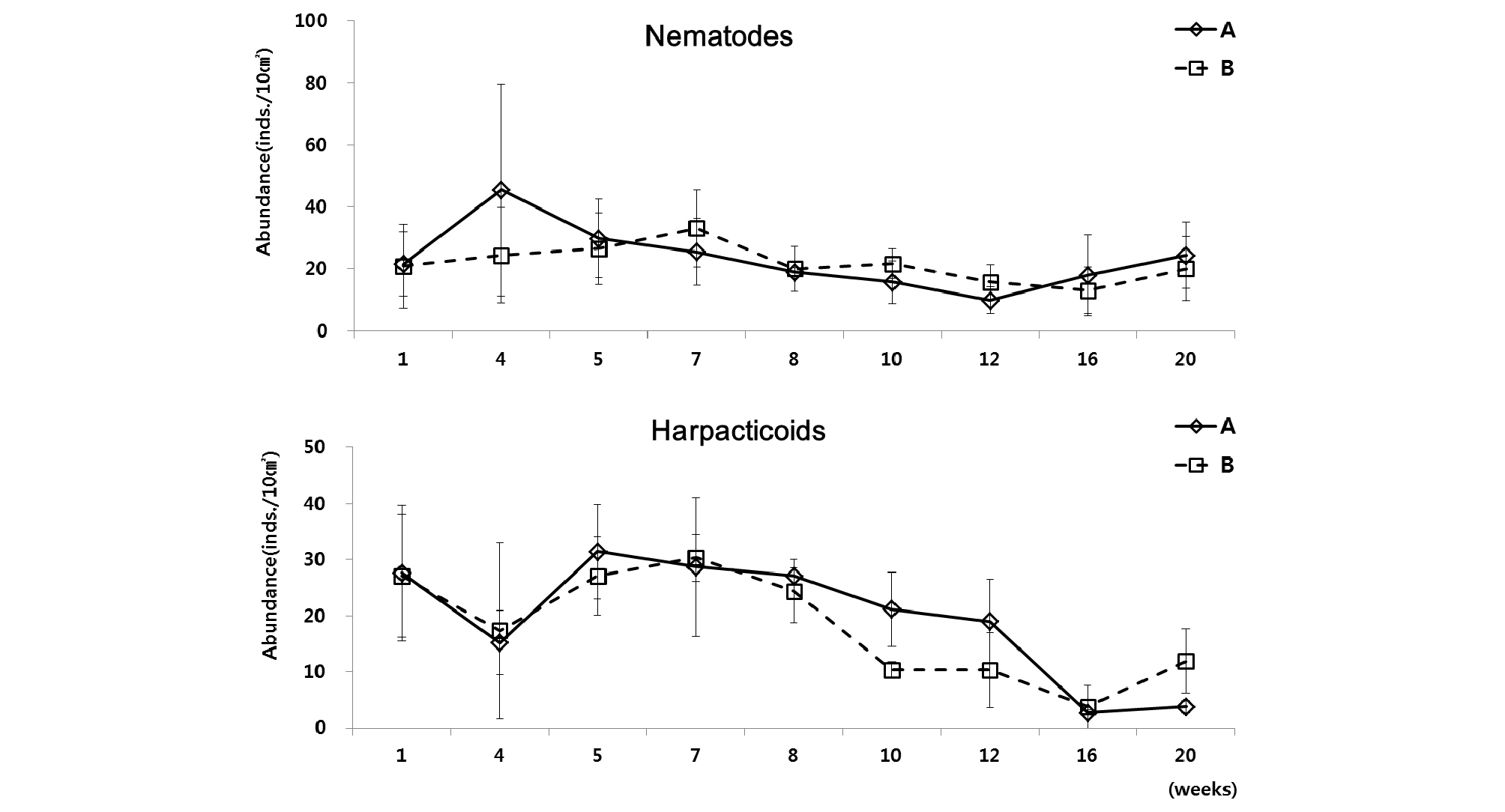
개체수가 가장 높은 선충류 및 저서성 요각류를 살펴보면, 수조 A와 B 모두 유사한 변화 양상을 보였다(Fig. 5). 선충류는 배양 초기인 배양 5주까지 최고 서식밀도가 수조 A에서 46 개체/10 ㎝², 수조 B에서 33 개체/10 ㎝²로 급격한 증가를 보이고 그 후 배양 16 주까지 서식밀도가 점점 감소하여 최소 서식밀도가 수조 A에서 10 개체/10 ㎝², 수조 B에서 13 개체/10 ㎝²로 나타났다. 이후 배양 20주까지는 서식밀도가 서서히 증가하는 양상을 보였다. 차우점하는 저서성 요각류에서는, 선충류와는 다르게 초기 배양 4주차에 급격히 서식밀도가 감소한 후, 1주일 뒤인 5주차에 급격히 상승하여 초기 서식밀도 값과 유사하게 회복되는 결과를 나타냈다. 배양 5주 이후부터 다시 서서히 감소하여 배양 16주에는 수조 A에서 3 개체/10 ㎝², 수조 B에서 4 개체/10 ㎝²로 최저값을 보였다. 그 이후로는 선충류와 마찬가지로 다소 회복하는 경향을 나타냈다(Table 2).
3.2 Exp. 2 : 먹이 공급 유무에 따른 중형저서동물 군집 구조 변화
두 번째 실험(Exp. 2)의 배양 실험 결과, 배양 초기의 전체 서식밀도 값은 수조 A와 B가 반대되는 양상을 보였지만, 배양 3주 이후로는 두 수조 내의 서식밀도 변화 양상은 서서히 감소하는 같은 결과를 보였다(Fig. 6). 배양 1 주 후, 자연 상태와 유사한 환경으로 배양한 수조 A에서는 290 개체/10 ㎝²로 급격하게 감소하였고, 먹이를 공급해준 수조 B는 768 개체/10 ㎝²로 급격하게 증가하는 서식밀도 값을 나타냈다. 그리고 배양 3주 후에 수조 A는 초기 서식밀도 값과 유사한 값까지 서식밀도가 회복되었고, 수조 B는 790 개체/10 ㎝²로 가장 높은 값을 나타낸 후, 배양 23주까지 서서히 감소되는 경향을 보였다(Table 3). 수조 A와 B의 중형저서동물 서식밀도는 먹이공급 유무에 따라 유의한 차이를 보였다(P<0.05)(Table 4).
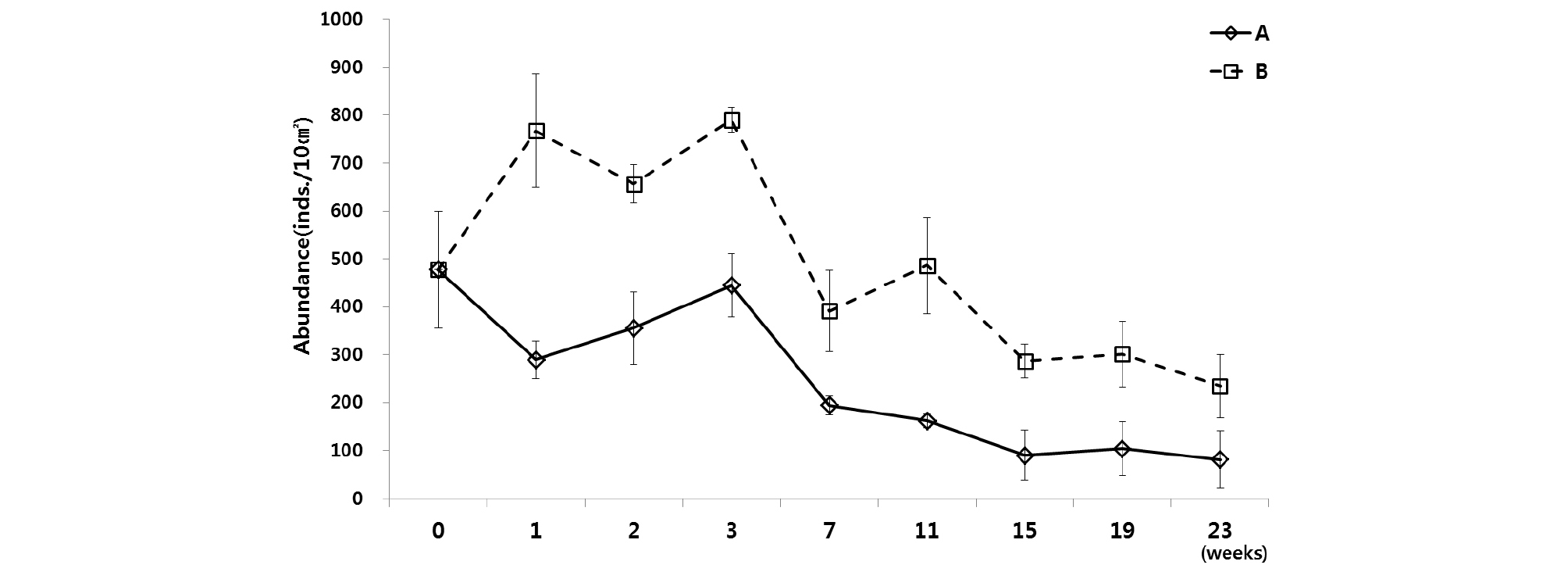
Table 3. Total density of meiofauna at experiment 2 (A: Light and no-food. B: Lingt and food containing algae and bacteria)
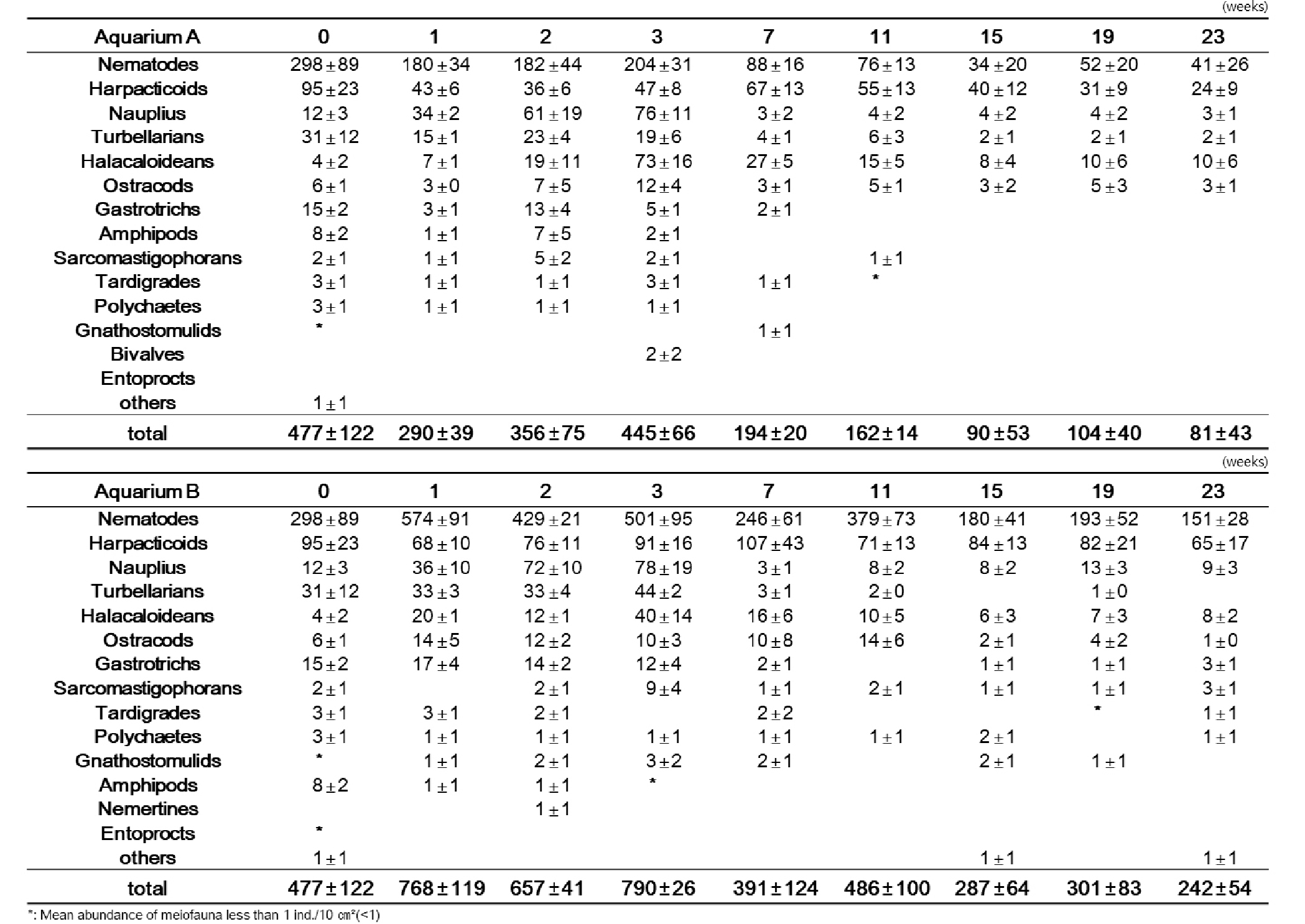 |
Table 4. Summary results of ANOVA analysis in experiment 2, 3
| Analysis | Total meiofauna density | |||
| F-value | P-value | |||
| Exp. 2 | One-way ANOVA | Food | 6.95 | <0.012 |
| Exp. 3 | Two-way ANOVA | Food | 11.364 | <0.001 |
| Light | 51.322 | <0.000 | ||
| Food*Light | 10.783 | <0.002 | ||
우점분류군인 선충류와 저서성 요각류는 서식밀도 변화에서 매우 다른 양상을 보였다. 선충류는 배양 3주 후, 저서성 요각류는 배양 7주 이후로는 서서히 감소하는 양상을 나타냈다(Fig. 7). 선충류의 경우, 전체 중형저서동물 서식밀도 변화와 유사한 변화를 보였다. 배양 1주 후에 수조 A에서 180 개체/10 ㎝²로 급격하게 감소하였고, 수조 B에서는 574 개체/10 ㎝²로 급증하였다가, 배양 3주부터 23주까지 서식밀도 값이 서서히 감소하는 양상을 보였다. 선충류 다음으로 우점한 분류군인 저서성 요각류의 경우, 배양 초기인 배양 1주에 서식밀도 값이 감소하였다가, 배양 7주에 수조 A에서 67 개체/10 ㎝², 수조 B에서 107 개체/10 ㎝²로 서서히 증가하였다. 그 이후로 배양 23주까지는 다시 서서히 감소하는 경향을 나타냈다(Table 3). 총 출현 분류군수는 수조 A에서 평균 9개로 나타났고 수조 B에서는 평균 11개로 나타났다(Table 3).
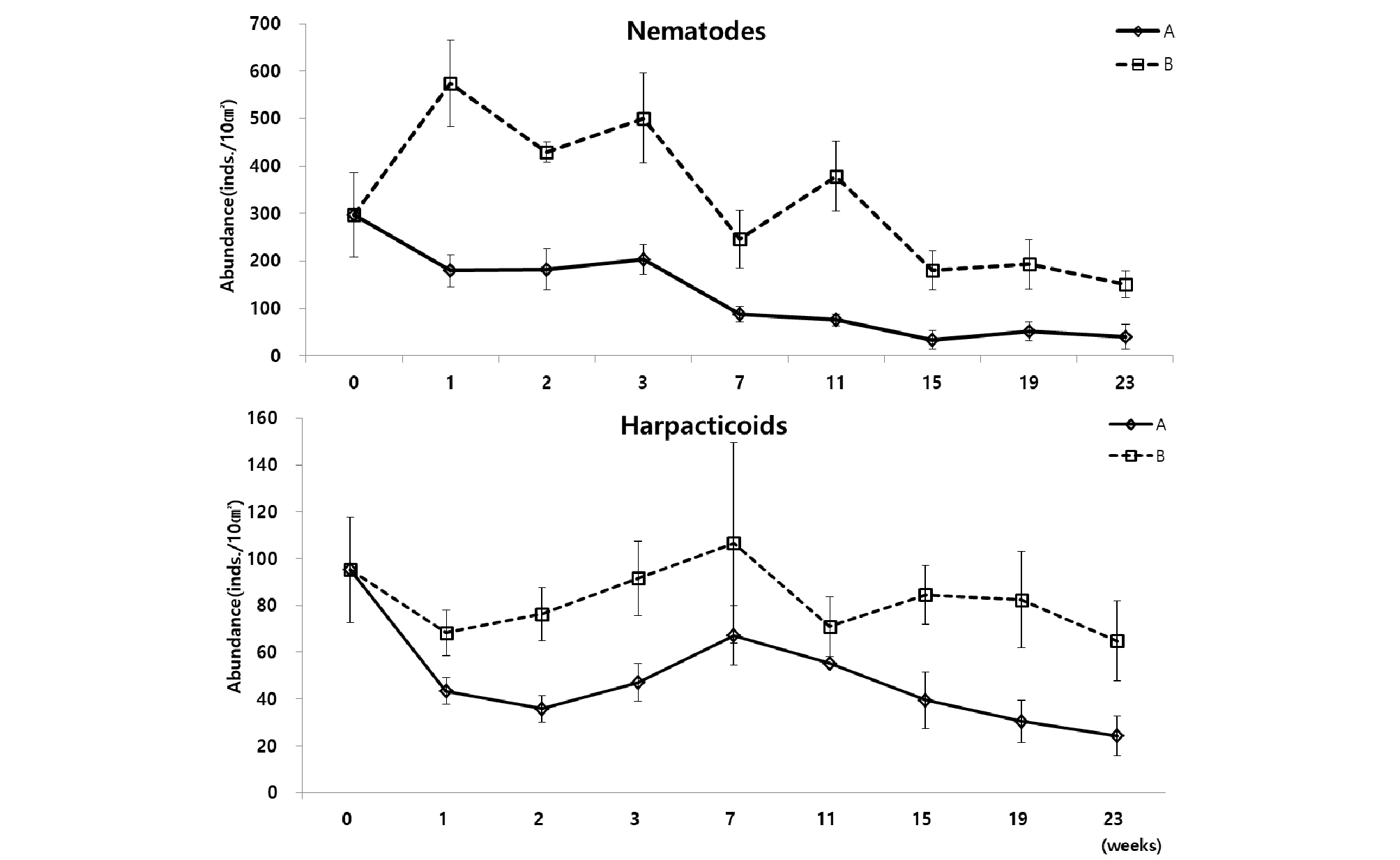
3.3 Exp. 3 : 먹이유무 및 광조건에 따른 군집 구조 변화
최종 실험(Exp. 3)의 수조 A~D 중에서 각 시기를 통틀어 서식밀도가 가장 높게 나타난 것은 수조 D로 나타났다. 다음으로 높게 나타난 것은 수조 C이다. 반면에 각 시기에 있어 전체적으로 가장 서식밀도가 낮게 나타난 것은 수조 A이다(Fig. 8). 가장 서식밀도가 높게 유지된 수조 D는 빛과 먹이 공급이 이뤄졌으며, 초기 먹이 공급 후, 8주에 걸쳐 서식밀도가 점차 증가하는 경향을 보이다가, 다시 배양 16주 후에 342 개체/10 ㎝²까지 서서히 감소하였고, 그 이후에는 배양 초기의 서식 개체수로 회복되며 안정된 모습을 보여주었다. 두 번째로 서식밀도가 높게 유지된 수조는 암배양 조건에서 먹이공급을 해준 수조 B로 나타났다. 수조 B는 배양 4주 후 서식밀도가 443 개체/10 ㎝²으로 증가하였다가 다시 서서히 감소하는 경향을 보였으며 배양 24주 후의 서식밀도는 196 개체/10 ㎝²로 초기 서식밀도의 55%를 유지하였다. 다음으로 서식밀도의 유지가 잘 되었던 수조 A는 암배양 조건에서 먹이를 공급하지 않은 조건이며, 서식밀도는 16주까지 서서히 감소하였다가 소폭 증가 후, 다시 감소하여 배양 24주 후에는 88 개체/10 ㎝²로 초기 개체수에 비하여 약 75%이 감소된 비율로 나타났다. 각 수조 배양의 조건에서 서식밀도의 가장 급격한 감소를 보인 수조 C는 12시간 주기의 광조건에서 먹이 공급을 하지 않은 조건이며, 배양 초기에 서식밀도의 급격한 감소 후, 소폭의 증가와 감소가 있었고 배양 24주 후에는 75 개체/10 ㎝²로 서식밀도의 약 20%의 생물만이 생존하는 결과를 나타내었다(Fig. 9, Table 5). 명배양과 암배양의 광조건과 먹이 공급의 유무에 따른 서식밀도는 각 조건에 따라 유의한 차이가 나타났다(P<0.05)(Table 4).
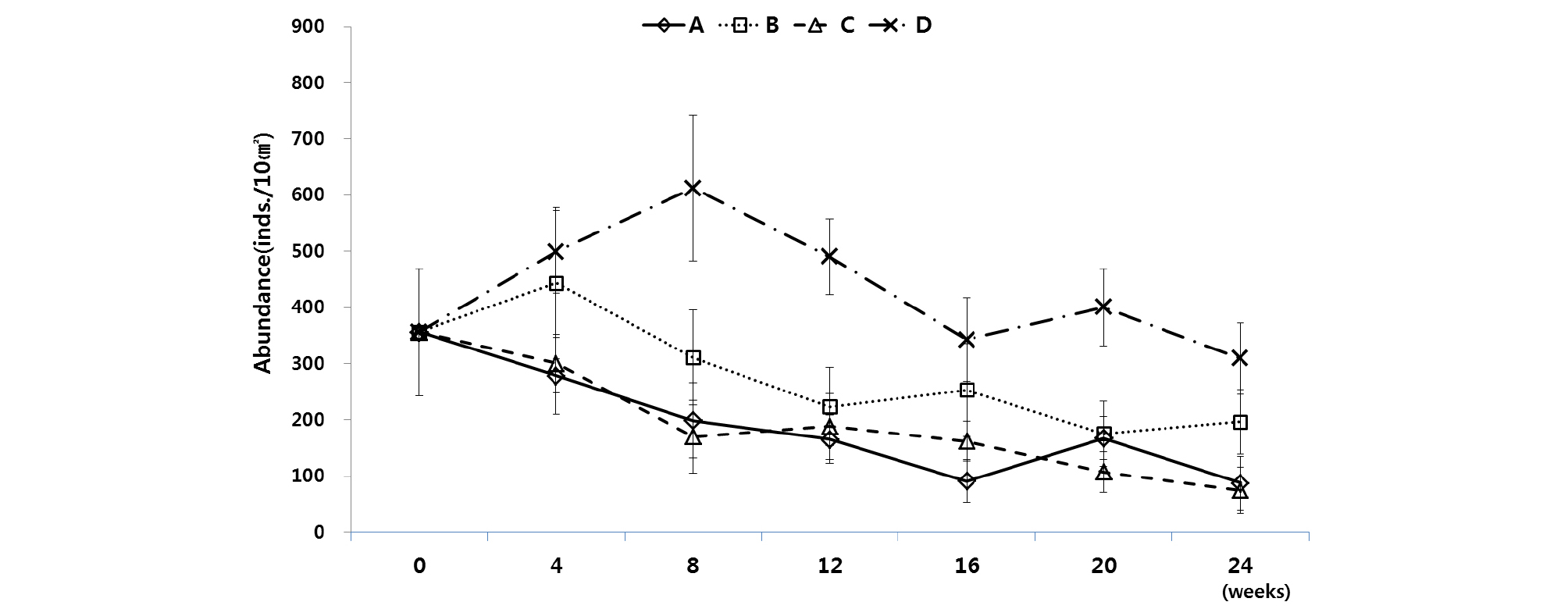
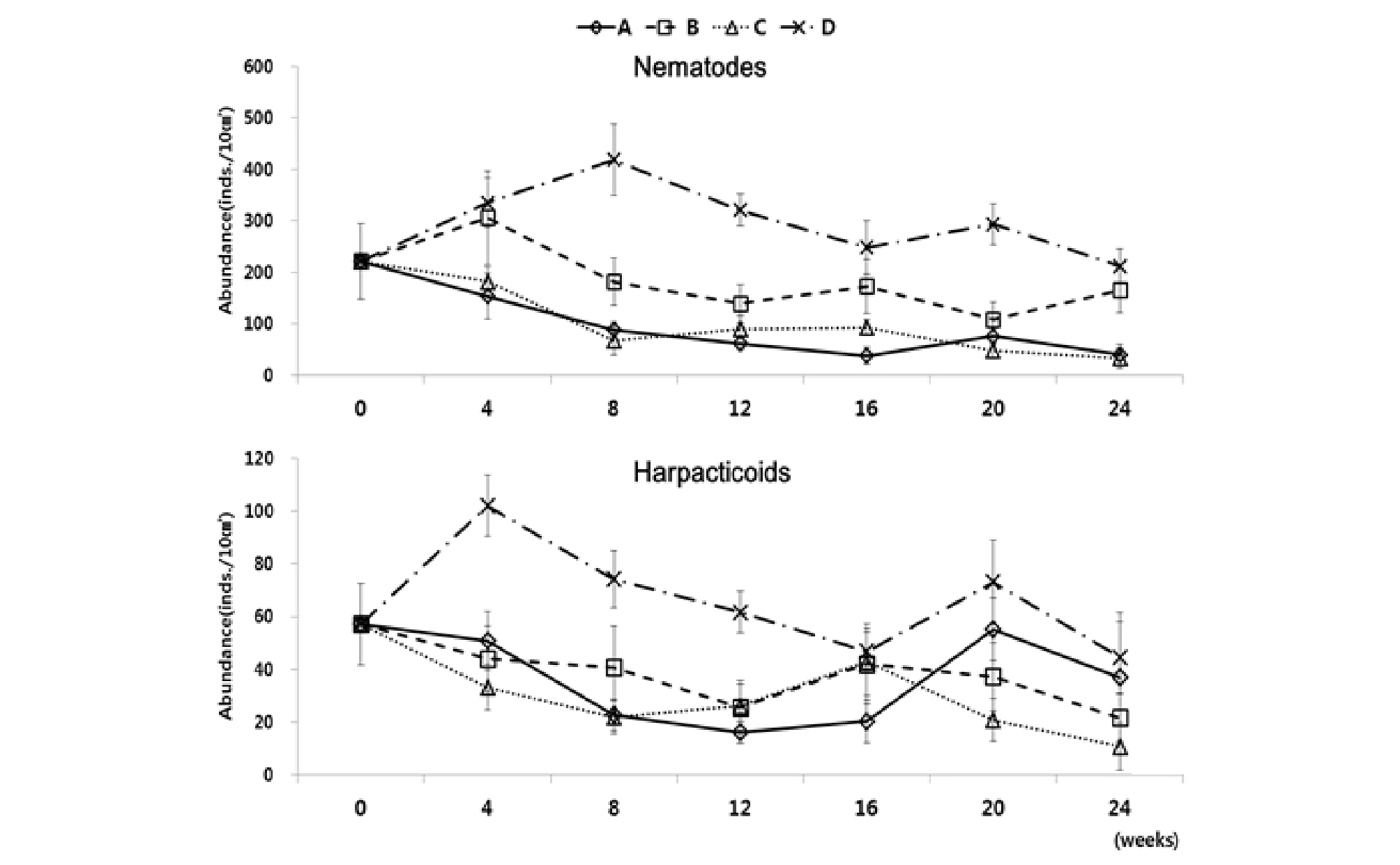
Table 5. Total density of meiofauna at experiment 3 (A: No-light and no-food, B: No-light and food containing diatoms and bacteria, C: Light and no-food, D: Light and food containing diatoms and bacteria)
 |
가장 우점하는 분류군인 선충류와 저서성 요각류의 서식밀도 변동을 살펴보면(Fig. 9), 먹이를 공급해 준 수조 B와 수조 D의 경우 배양 직후 서식밀도가 수조 B에서 306 개체/10 ㎝², 수조 D에서 335 개체/10 ㎝²까지 증가하였다가 다시 서서히 감소한 뒤, 배양 24 주 후에 수조 B에서 165 개체/10 ㎝², 수조 D에서 212 개체/10 ㎝²까지로, 초기 서식밀도 값과 유사한 값을 보였다. 반면 먹이를 공급해주지 않은 수조 A와 C의 경우, 배양 초기인 8주까지 서서히 감소하여 수조 A에서 88 개체/10 ㎝², 수조 C에서 68 개체/10 ㎝²의 서식밀도를 보이고 그 이후 작은 폭의 증가와 감소를 보이다가 배양 24주 후에는 수조 A에서 41 개체/10 ㎝², 수조 C에서 34 개체/10 ㎝²로 매우 낮은 값까지 감소하는 결과를 나타냈다. 저서성 요각류의 경우, 선충류와는 다소 다른 양상을 나타냈다. 수조 A, B, C에서는 배양 직후 서식밀도가 계속 감소하다가 배양 16주 이후로 다시 증가하여 수조 A에서 55 개체/10 ㎝², 수조 B에서 42 개체/10 ㎝², 수조 C에서 43 개체/10 ㎝²의 값을 나타내었다. 배양 24주 후에는 수조 A에서 37 개체/10 ㎝², 수조 B에서 22 개체/10 ㎝², 수조 C에서 11 개체/10 ㎝²로 다시 감소하는 결과를 나타냈다(Table 5).
총 출현 분류군수는 수조 A와 B에서 평균 9개, 수조 C에서는 평균 10개로 나타났으며 수조 D에서는 평균 12개로 가장 많은 출현분류군 수를 나타냈다(Table 5).
4. 고 찰
자연 상태 조건에서의 실험(Exp. 1) 결과, 중형저서동물의 서식밀도는 대부분 선충류와 저서성 요각류의 서식밀도에 영향을 받았으며, 전체 서식밀도의 변화는 가장 우점하는 선충류의 서식밀도 변화와 거의 유사한 양상을 보였고, 이는 자연 상태에서도 같은 경향을 나타낸다(Braeckman et al., 2013).
먹이 공급 유무에 따른 실험(Exp. 2)에서는 먹이공급을 하지 않은 수조 A에서 초기 서식밀도가 감소하였지만, 먹이로 박테리아와 조류를 공급한 실험구인 수조 B의 서식밀도는 급격히 증가하였다. 그 이후에도 수조 B에서의 서식밀도는 비교적 높게 유지되는 결과를 보였고, 첫 번째 실험에서처럼 선충류의 서식밀도 변화는 전체 서식밀도 변화와 유사한 양상을 나타내었다.
각 배양 조건의 다양성 분석 결과, 풍부도(d)는 세 번째 실험의 수조 D에서 가장 높았고, 균등도(J’)와 분류군 다양도 지수{H’(loge)}는 세 번째 실험의 수조 C에서 가장 높게 나타났다(Table 6).
Table 6. Analysis of diversity at each Aquarium (D=Richness, J'=Evenness, H''loge=diversity)
| Aquarium | D | J' | H'loge | |
| Exp.1 | A | 1.4 | 0.5 | 0.9 |
| B | 1.4 | 0.6 | 1.0 | |
| Exp.2 | A | 1.1 | 0.7 | 1.3 |
| B | 1.6 | 0.5 | 1.1 | |
| Exp.3 | A | 0.9 | 0.7 | 1.1 |
| B | 0.9 | 0.3 | 0.6 | |
| C | 1.2 | 0.8 | 1.5 | |
| D | 1.7 | 0.4 | 1.1 |
먹이 공급 및 광조건 유무에 따른 실험(Exp. 3)에는 두 조건을 동시에 적용하여 실험을 수행하였다. 암배양 조건은 빛으로 인한 조류의 급속한 성장을 방지하기 위하여 설정하였다. 선충류의 서식밀도 변화는 전체 중형저서동물 서식밀도와 유사한 양상을 나타내었다. 암배양 조건에서의 저서성 요각류의 서식밀도 변화와 전체서식밀도 변화 양상이 일치하지 않는 이유는 저서성 요각류가 다른 중형저서동물 분류군에 비하여 오염이나 주변 환경에 상대적으로 더 민감하고(McLachlan, 1978; Lee et al., 2001; Wetzel et al., 2001), 인위적 변화에 서식밀도가 영향을 많이 받는 분류군(Lee et al., 2001)이기 때문인 것으로 추측된다. 또한 광원의 조도에 따라 요각류의 섭식률과 개체발생기간에 차이가 있다고 알려져 있는데(Li et al., 2008; Lee et al., 2011), 본 연구에서 배양된 요각류 종에 배양수조에 이용된 광원의 조도가 어떤 영향을 미쳤는지를 정확하게 파악하기 위해서는 향후 개체군 배양이 수행되어야 할 필요성이 있다.
직접 설계 제작된 수조에는 실험실 내에서 중형저서동물의 군집 규모의 배양을 위한 장비들을 설치하였다. 주요 장비 중, 산소공급기는 배양수질악화 및 산소부족으로 인한 영향을 최소화하기 위하여 일반 관상용 수조에 들어가는 것 보다 높은 효율의 설계를 하였다. 이는 일반적으로 해양 퇴적물 내의 중형저서동물 생체량은 대형저서동물(macrofauna)에 비하여 매우 낮지만(Giere, 2009), 산소 소비량은 대형저서동물의 다섯 배 이상을 소비하기 때문이다(Gerlach, 1971). 또한 퇴적물의 표층에서부터 깊이가 깊어질수록 유기물질의 순환이 원활하게 이뤄지지 않아 중형저서동물 및 그들의 먹이원으로 이용되는 박테리아의 생존률에도 부정적인 영향을 미치는데(Heip et al., 2001; Thistle, 2001; Rowe et al., 2003; Rex et al., 2006; Gontikaki et al., 2011; Zeppilli et al., 2015), 이와 같은 현상을 방지하기 위하여 수조의 하단부에 다공성인 구조물을 설치하였다. 그 위에 퇴적물이 다공성 구조물을 통과하거나 끼어서 해수 흐름을 막지 않도록 섬유물질을 배치하였다, 수조 내의 해수는 퇴적물 표층이 교란되지 않도록 상층에서부터 여러 종류의 여과재로 이루어진 필터를 통하여 여과시킨 후에 퇴적물 하층에서 퇴적물의 표층으로 이동시키는 방식을 이용하여 순환시켰다. 이러한 수조의 설비로 인해 실험 수행 중에 퇴적물의 부패가 방지되어 중형저서동물의 서식밀도가 장기간 유지될 수 있었고, 그로 인하여 20~24주의 실험실 내 배양을 지속할 수 있다고 판단되어진다.
자연 상태에서 만리포 조간대 중부의 중형저서동물 군집의 서식밀도는 2008년에 47~880 개체/10 ㎝², 2009년에 10~708 개체/10 ㎝², 2010년에 59~754 개체/10 ㎝², 2011년에 14~391 개체/10 ㎝², 2012년에 38~350 개체/10 ㎝²로 계절에 따라 서식밀도 차이가 매우 큰 결과를 나타내었는데(Kang et al., 2016), 본 연구의 결과처럼 실험실 내에서 중형저서동물 군집의 서식밀도를 계절과 상관없이 일정하게 유지할 수 있다면 중형저서동물을 이용한 실험을 수행할 때, 매번 현장에서 시료를 조달하는 방법에 비하여 더욱 효율적인 연구를 할 수 있을 것이라고 생각된다.
배양 실험을 종료할 때의 기준으로 출현분류군수를 살펴보면, 두 번째 실험의 먹이를 공급하지 않은 수조 A에서는 최종 출현분류군수가 6개였고, 먹이를 공급한 수조 B의 경우 이보다 더 많은 10개를 나타내었다. 세 번째 실험의 암배양에서도 먹이를 공급하지 않은 수조 A는 5개, 먹이를 공급한 수조 B에서는 6개로 작은 차이이지만 출현분류군수에서 차이를 보였고, 명배양에서 먹이를 공급하지 않은 수조 C에서의 최종 출현분류군수는 6개였고, 먹이를 공급한 수조 D에서는 11개의 값을 나타내었다.
이러한 결과로 미루어보아, 먹이의 공급은 서식밀도 뿐만 아니라 출현분류군수에도 영향을 미친다고 여겨지며 먹이를 공급하지 않은 조건에 비하여 먹이를 공급해 준 조건이 중형저서동물 군집 규모의 배양에서 초기 서식밀도와 출현분류군수를 유지하는데 더욱 적합한 조건이라고 여겨진다. 또한 암배양보다는 자연조건과 유사한 주기의 광조건을 설정하는 것이 상대적으로 더 높은 서식밀도와 출현분류군수를 나타내는 결과를 보였다. 종합적으로, 중형저서동물의 군집 규모의 배양에서는 명조건과 먹이를 공급해주는 조건이 배양을 유지하는데 유리한 조건이라고 생각된다.
앞에서 여러 조건에 따라 다양한 결과들 각각의 의미에 대해서 설명을 하였다. 본 연구를 통해 나타난 그 특징의 요점을 간략히 나열하면, 중형저서동물의 배양 유지에 효율적인 항목과 조건은 가) 수조의 설계 시 퇴적물의 부패를 방지하기 위한 구조물과 해수 순환 구조, 나) 먹이조건으로 박테리아와 저서성 규조류를 공급, 다) 12시간 주기의 광조건으로 나타났다.
향후 그 실험과 진행이 무척 어려울 것이라 예상되며, 보다 세밀하고 체계적인 계획이 요구될 것이다. 본 연구를 바탕으로 우점 출현분류군의 종조성 분석 등을 통한, 먹이 조건과 광조건 등이 각 분류군 및 개체군의 종 수준에 이르기까지 미치는 영향을 파악하는 세밀한 연구가 필요하다고 생각된다. 더불어 인위적인 환경에서의 적응력이 높은 분류군에 관한 분석 또는 환경 변화에 민감하여 환경 오염 등의 지표종으로써 이용가능한 분류군 등이 파악된다면 향후 한층 더 중형저서동물에 관한 다양한 연구에 활용 가능할 것이라 생각된다. 이러한 연구를 통해 중형저서동물 군집 규모의 배양을 효율적으로 유지 가능하게 되면, 중형저서동물을 이용한 실험 때마다 직접 현장에 나가 시료를 채집하는 시간적·경제적 소모를 현격히 줄일 수 있으며 연구의 효율성을 높이게 될 것이라 생각한다.
