

1. 서 론
2. 재료 및 방법
2.1 조사장소 및 왕거머리말 이식
2.2 환경요인 측정
2.3 이식잘피(transplants) 모니터링
2.4 광합성 특성 측정
2.5 통계분석
3. 결 과
3.1 환경요인
3.2 왕거머리말의 생육밀도, 성장률, 형태학적 특성
3.3 광합성 특성
4. 고 찰
1. 서 론
잘피(seagrass)는 연안 및 하구생태계에 분포하는 유일한 고등현화식물(marine angiosperm)로서, 높은 성장률을 바탕으로 다양한 생태계서비스(ecosystem services)들을 제공해준다(Costanza et al., 2014; Nordlund et al., 2016; Ruiz-Frau et al., 2017). 잘피생육지는 다양한 해양생물들에게 직·간접적인 먹이원과 어류 자원들의 산란장 및 서식처를 제공하므로 연안 생태계에서 매우 중요한 역할을 담당하고 있다(Kritzer et al., 2016; Whippo et al., 2018; Unsworth et al., 2019). 또한, 유속 및 파랑에너지 감소를 통해 해저퇴적물을 안정화시키고, 과도하게 유입되는 무기영양염류나 오염물질을 흡수·제거하는 뛰어난 생태계 정화능력을 가지고 있다(Choi et al., 2016; Sandoval-Gil et al., 2016; Potouroglou et al., 2017). 잘피는 광합성을 통해 온실가스 중의 하나인 이산화탄소를 흡수하는 능력이 뛰어나기 때문에, 최근에는 블루카본 생태계(blue carbon ecosystem)로써 지구온난화를 완화시킬 수 있는 중요한 해양생물자원으로 인식되고 있다(Mcleod et al., 2011; Fourqurean et al., 2012; Duarte et al., 2013). 그러나 전 세계적으로 잘피생육지의 면적은 자연적 요인(태풍, 기후변화, 질병, 섭식 등) 및 인위적 요인(준설, 매립, 건설, 어업활동 등)으로 인해 급격히 감소하고 있으며, 우리나라의 잘피생육지 또한 50% 이상 훼손되거나 사라졌다고 추정된다(Waycott et al., 2009; Lee et al., 2018; Unsworth et al., 2018).
잘피의 생장은 해양의 다양한 환경요인에 의해 영향을 받는데, 주로 수중광량, 수온 및 무기영양염에 의해 조절되는 것으로 알려져 있다(Lee et al., 2007b). 또한 생육조건이 변하거나 새로운 환경에 노출되면 잘피는 다양한 환경에 적응하기 위해 생리생태학적 변화가 나타난다(Collier et al., 2007; Cabaço et al., 2009; Park et al., 2016b). 잘피는 생장과 생존을 위해 광합성에 필요한 충분한 수중광량이 필요하며, 수중광량의 변화는 잘피의 분포와 생장에 중요한 요인으로 고려되고 있다(Ralph et al., 2007). 수온은 잘피의 계절적인 생장을 조절하는 주요 요인으로, 계절이 뚜렷한 온대 해역의 잘피는 수온이 증가하는 봄과 초여름에 활발한 생장을 보이고, 가을과 겨울에 생장이 감소한다(Lee et al., 2007b). 국내 연안에 분포하는 거머리말은 약 15-20℃, 게바다말은 약 14℃, 왕거머리말은 약 16-19℃에서 최대의 성장률을 보이며, 20℃ 이상의 여름철 고수온에서 극심한 생장저해를 나타낸다(Lee et al., 2005; Park and Lee, 2009; Park et al., 2016b). 또한 잘피는 잎과 뿌리 모두를 통해 무기영양염을 흡수할 수 있기 때문에, 해수 및 퇴적물 공극수의 무기영양염 농도는 잘피의 생장을 조절한다. 잘피생육지 내 무기영양염 농도가 낮으면 잘피의 생장이 저해되고, 반면에 무기영양염 농도가 너무 높으면 부착조류 및 플랑크톤의 번성을 야기하여 수중광량 감소 또는 산소결핍 등을 통해 잘피의 생장을 저해시킨다.
잘피는 남극을 제외한 전 세계 연안에 약 60-70여 종이 널리 분포하고 있다(Short et al., 2007). 그중 우리나라에는 5종의 거머리말 속[거머리말(Zostera marina), 포기거머리말(Z. caespitosa), 수거머리말(Z. caulescens), 애기거머리말(Z. japonica), 왕거머리말(Z. asiatica)], 2종의 새우말속[새우말(Phyllospadix iwatensis), 게바다말(P. japonicus)], 그리고 아열대 생장 특성을 가진 해호말(Halophila nipponica)과 기수종인 줄말(Ruppia maritima)까지 총 9종의 잘피가 분포하고 있다(Lee et al., 2018). 이 중 왕거머리말은 다른 종들과는 달리 우리나라 동해안에만 분포한다(Park et al., 2016a; Lee et al., 2018). 왕거머리말은 우리나라 동해안과 일본 북부의 일부 해역에만 분포하며, 러시아 동쪽과 중국 발해만, 미국 서부 캘리포니아 연안에서도 작은 면적이 일부 보고되었다(Phillips and Echeverria, 1990; Lee et al., 1999; Green et al., 2003; Wyllie-Echeverria and Ackerman, 2003; Tanaka et al., 2009; Shi et al., 2010). 이러한 제한적인 분포 뿐만 아니라 생육지 면적이 현저하게 감소하고 있어, 왕거머리말은 국제자연보전연맹 적색목록(IUCN Red list)에 위기근접종(NT: Near Threatened)으로 등재되어 있으며, 우리나라에서는 “해양생태계의 보전 및 관리에 관한 법률”에 의해 해양보호생물(marine organisms under protection)로 지정되어 있다. 왕거머리말의 연구는 주로 분포와 형태에 대해서만 수행되어 왔으며, 최근 우리나라 동해안에서 생장 특성에 대한 연구가 수행되었다(Lee et al., 1999; Watanabe et al., 2005; Tanaka et al., 2009; Park et al., 2016a).
우리나라의 동해안은 대륙붕의 발달이 미약하며, 수심이 깊고 해안선이 매우 단조로우며 파랑에너지가 높은 것이 특징이다. 따라서 동해안의 잘피는 암반에 부착하여 생육하는 새우말속 잘피들이 우점하고 있으며, 거머리말속 잘피종들은 작은 항구나 석호와 같은 폐쇄된 해역에 작은 패치를 형성하여 분포한다(Lee et al., 2018). 우리나라 연안에 분포하는 대부분의 거머리말속 잘피종들은 비교적 얕은 수심의 잔잔한 해역에 분포하는 것과는 달리 왕거머리말은 파랑에너지가 높고 개방된 동해 연안의 깊은 수심에 분포한다(Lee et al., 1999; Park et al., 2016a; Lee et al., 2018). 미국과 일본의 왕거머리말은 각각 수심 5-17 m와 수심 4-10.8 m에 분포하여 깊은 수심 뿐만 아니라 비교적 얕은 수심에도 분포하고 있으나(Phillips and Echeverria, 1990; Tanaka et al., 2009), 우리 동해 연안의 왕거머리말은 8-15 m의 깊은 수심에 한정되어 분포한다. 이에 본 연구는 우리나라 동해안의 왕거머리말이 깊은 수심에 한정되어 분포하는 이유를 알아보고자 하였다. 이를 위해 깊은 수심의 왕거머리말을 얕은 수심으로 인위적으로 이식한 후 생리생태학적 변화를 자연생육지의 개체와 비교하여 파악하였다.
2. 재료 및 방법
2.1 조사장소 및 왕거머리말 이식
본 연구는 우리나라 동해 연안의 울진군 죽변면 죽변항에서 남쪽으로 약 500 m 떨어진 곳에 분포하는 왕거머리말(37°03′05″N, 129°25′06″E)을 대상으로 2011년 10월부터 2012년 9월까지 수행되었다(Fig. 1). 왕거머리말은 수심 약 9-15 m에 분포하고 있으며, 소규모의 수거머리말 패치가 혼재되어 있다. 왕거머리말의 이식은 죽변항 서쪽 방파제 외부의 거머리말 생육지 인근의 수심 약 3 m 해역에서 진행되었다. 자연생육지(reference site) 및 이식 장소(transplant site)의 퇴적물은 모두 사질로 이루어졌다.
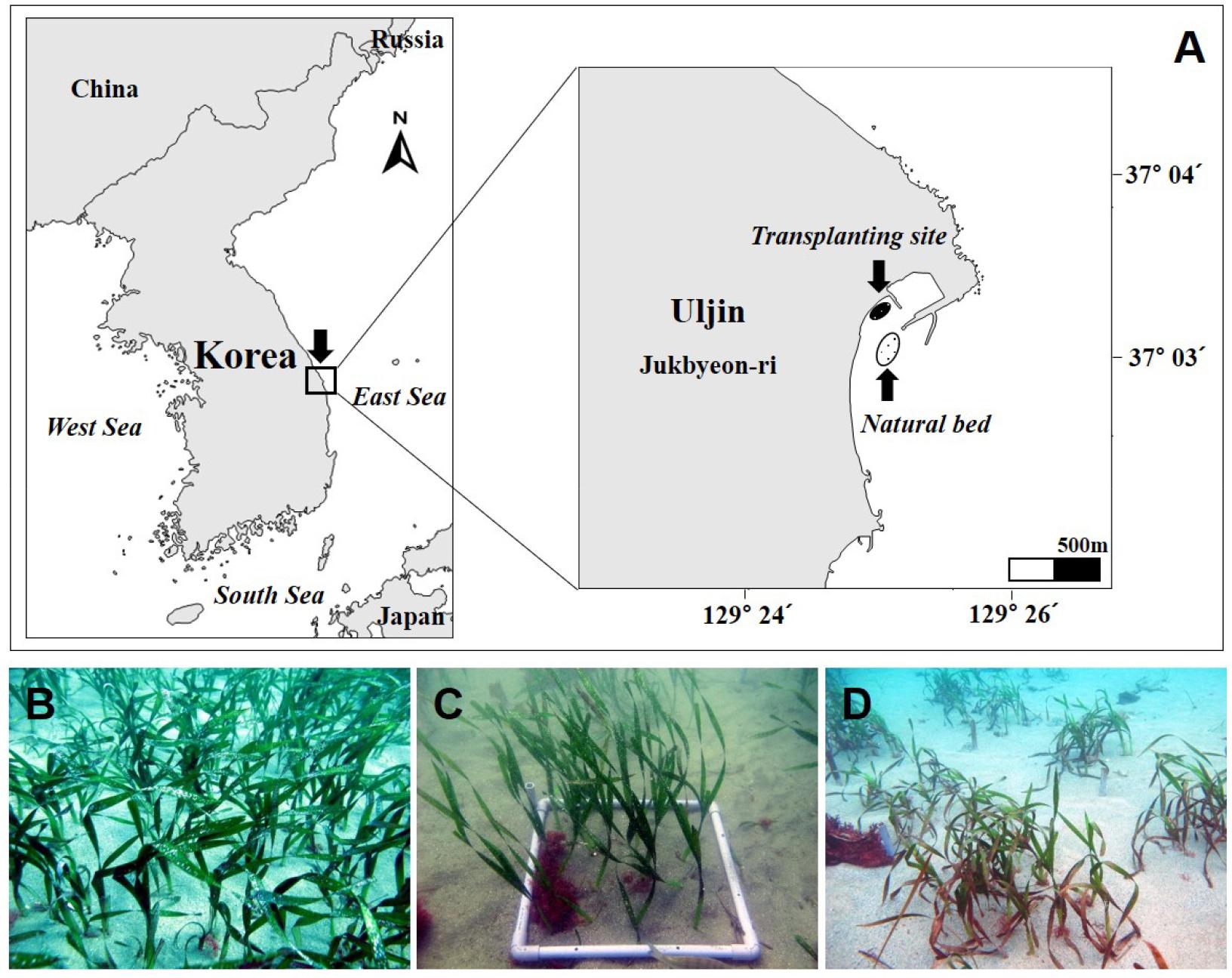
왕거머리말을 이식하기 위해 수심 9 m의 왕거머리말 자연생육지에서 개체들을 채취하여 자연생육지로부터 약 500 m 거리의 수심 3 m 지점에 왕거머리말을 이식하였다. 이식에 필요한 잘피 개체는 자연생육지의 훼손을 최소화하기 위해 자연생육지 모니터링 지점과 떨어진 곳에서 일정한 간격을 유지하며 채취하였다. 해수를 담은 플라스틱 용기에 채취한 개체를 넣어 즉시 이식 장소로 이동하여 이식하였다. 이식은 2011년 10월에 진행하였으며, 여러 이식 방법 중 높은 생존율을 보이는 철사고정방법(staple method)을 사용하였다(Park et al., 2005). 이식개체로 지하경 마디가 6개 이상인 개체를 이용하였으며, 자연생육지와 유사한 생육 밀도로 이식하기 위해 0.5 × 0.5 m 방형구 내에 15개의 개체를 각각 하나씩 철사에 고정하여 이식하였다. 총 10개의 방형구(2.5 m2)에 150 개체의 왕거머리말을 이식하였다.
2.2 환경요인 측정
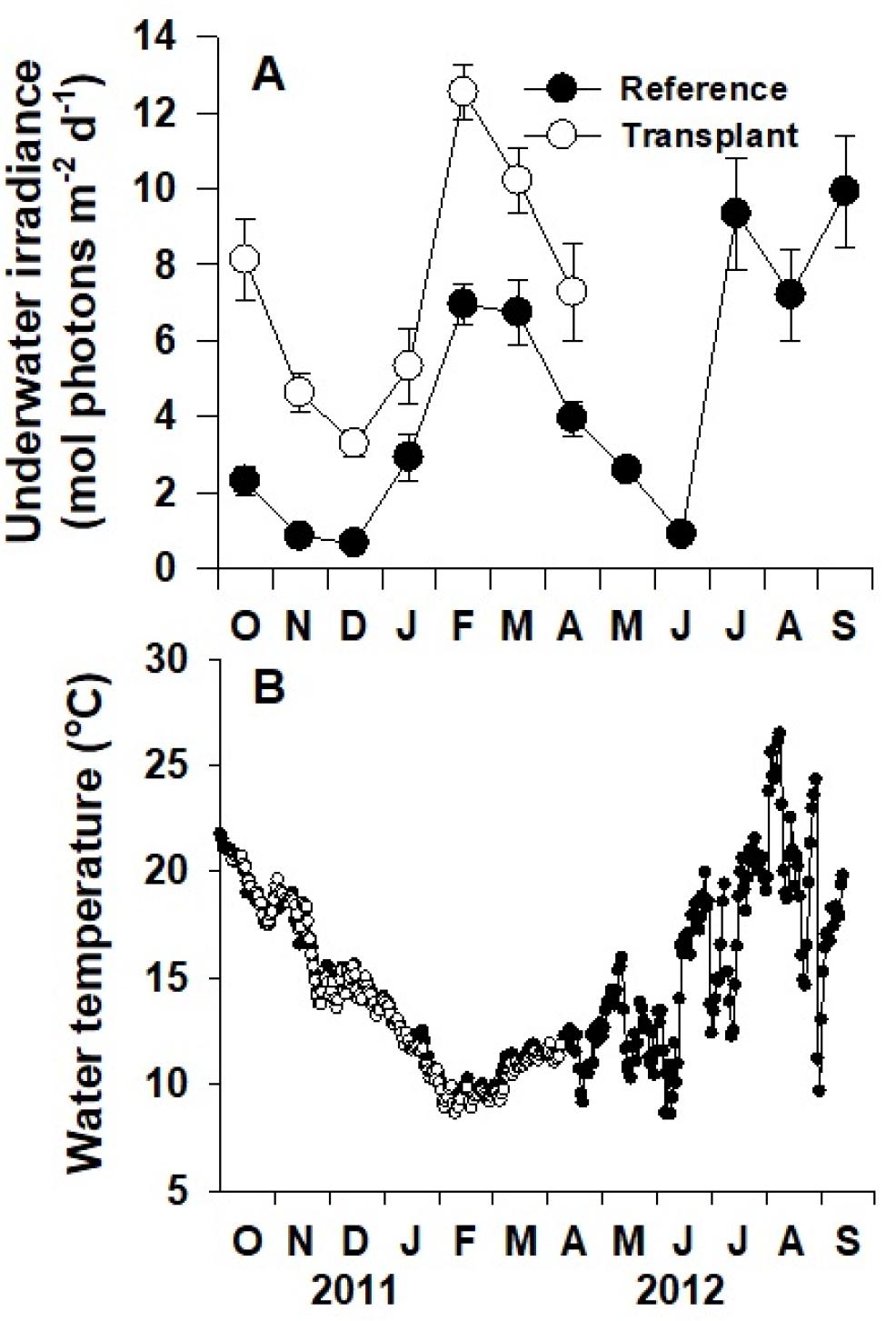
수중광량은 자연생육지와 이식 장소에서 왕거머리말의 잎이 위치한 수심에 Odyssey Photosynthetic Active Radiation (PAR) recorder (Dataflow Systems, Christchurch, New Zealand)를 설치하여 매 15분 간격으로 측정하였다. Odyssey PAR recorder를 통해 측정된 값은 동시간에 LI-193SA 센서와 LI-140 기록계(Li-Cor, Lincoln, NE, USA)를 이용하여 측정된 수치와의 회귀분석을 통하여 photon flux density (PFD, μmol photons m−2 s−1)로 변환하였으며, 하루 동안의 총 광량인 일일광량(mol photons m−2 d−1)으로 나타내었다. 수온은 각 조사장소에서 HOBO pendant® Temperature (Onset computer Corporation Bourne, MA, USA)센서를 설치하여 매 15분 간격으로 측정한 후 일일평균수온으로 나타내었다. 조사기간 중 강한 파랑에너지에 의해 Odyssey센서 및 HOBO센서를 2012년 중순 이후 소실하였고, 따라서 추후 통계분석은 2011년 10월부터 2012년 4월까지의 값을 사용하였다.
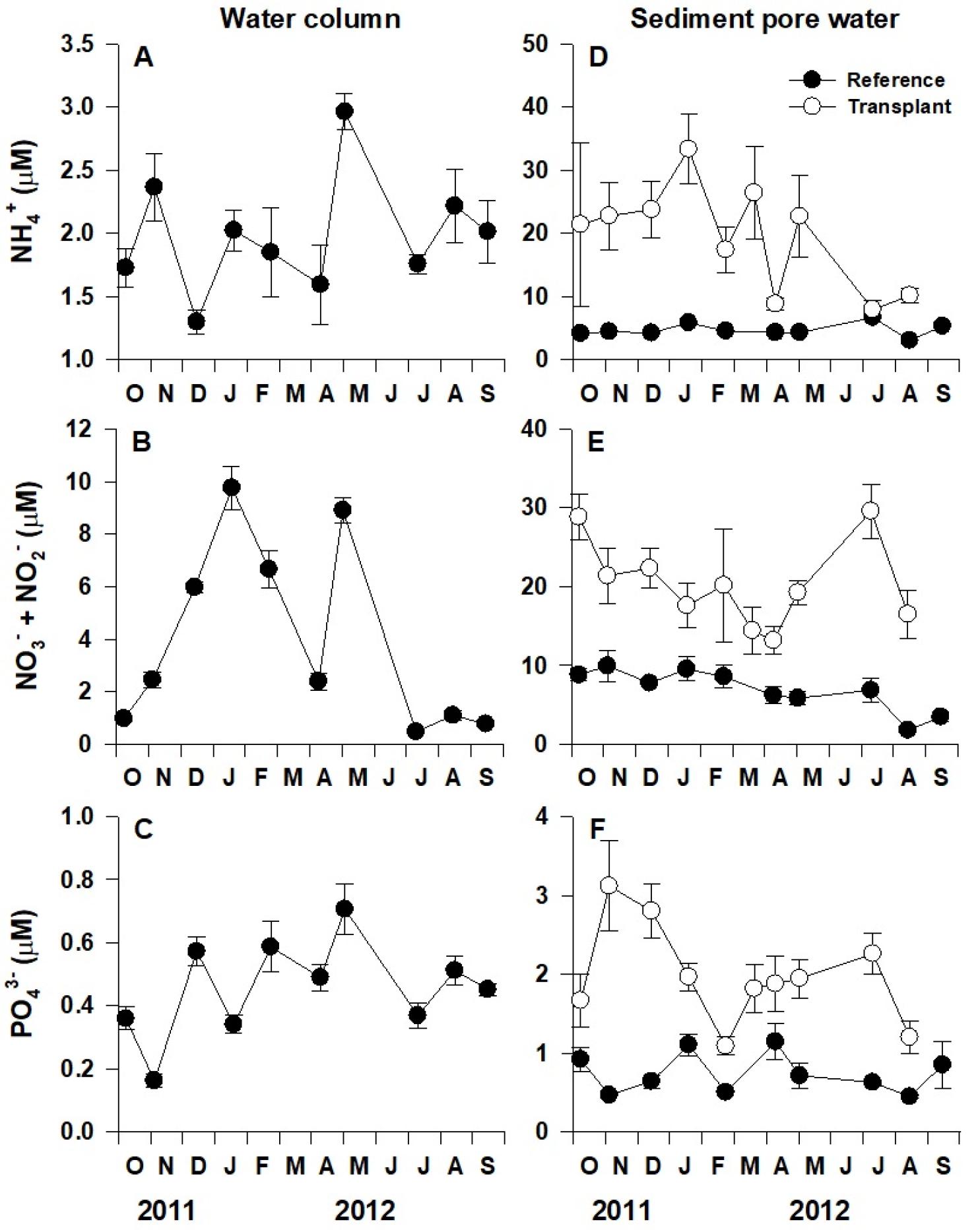
해수(water column)와 퇴적물 공극수(sediment porewater) 내 용존무기질소(NH4+, NO3−+NO2−)와 용존 무기인(PO43−)의 농도는 표준비색법(standard colorimetric techniques)을 이용하여 분석하였다(Parsons et al., 1984). 해수 시료(n=4)는 2011년 10월부터 2012년 9월까지 자연생육지에서 채취하였으며(2012년 3월, 6월 제외), 퇴적물 시료(n=4)는 2011년 10월부터 2012년 9월까지 자연생육지(2012년 3월, 6월 제외)와 이식 장소(2012년 6월, 9월 제외) 모두에서 채취하였다. 퇴적물 시료의 채취는 지름 5 cm, 길이 15 cm의 원형 아크릴코어를 이용하였으며, 이후 퇴적물을 원심분리(8000 rpm, 20 min)하여 상층부의 공극수를 채취하였다. NO3−+NO2− 농도는 구리로 코팅된 카드뮴 환원관을 이용하여 NO3−를 NO2−로 환원한 후 측정하였다.
2.3 이식잘피(transplants) 모니터링
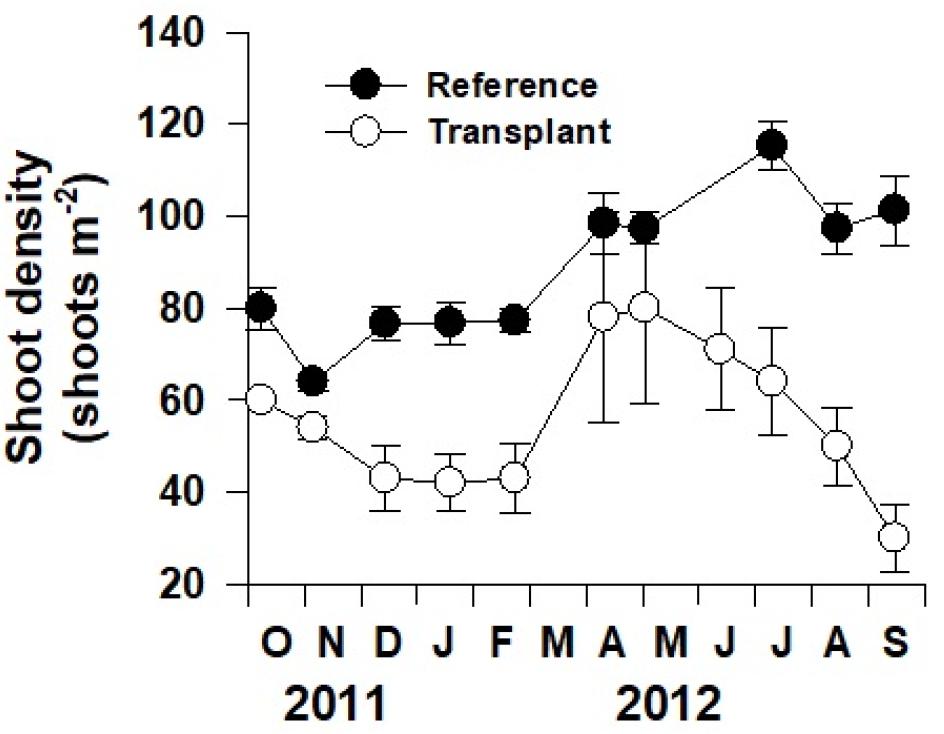
얕은 수심에 이식된 잘피의 정착과정을 파악하기 위하여 이식 후 매월 자연생육지와 이식 장소에서 잘피의 생육밀도(shoot density), 성장률(productivity) 및 형태적 특성(morphology)을 측정하였다. 생육밀도는 방형구(0.5 × 0.5 m) 내 왕거머리말의 개체수를 측정하여 단위면적당 생육밀도(shoots m−2)로 나타내었다. 왕거머리말 잎의 성장률은 modified blade marking technique를 이용하여 측정하였다(Kentula and McIntire, 1986; Lee et al., 2005). 매월 10개 내외의 잘피 개체를 임의로 선정하여 주사바늘을 이용해 엽초에 구멍을 뚫어 표시하였다. 한 달 후 구멍을 기준으로 새로 자란 부분(new leaf)과 기존 부분(old leaf)으로 분리하여 60℃에서 일정한 무게가 될 때까지 건조시킨 후 각각의 건중량을 측정하였으며, 개체당 일일 잎 성장률(mg DW shoot−1 day−1)으로 계산하였다. 또한, 단위면적당 잎 성장률(g DW m−2 day−1)은 개체당 잎 성장률에 생육밀도를 곱하여 계산하였다. 형태적 특성은 측정을 위해 회수한 개체의 전체 길이(cm)와 잎 너비(mm) 등을 측정하였다.
2.4 광합성 특성 측정
자연생육지와 이식 장소에서 왕거머리말의 광합성 특성을 비교하기 위해 엽록소형광측정기(Diving-PAM, Walz, Germany)를 사용하였다. 엽록소 형광 측정법(Pulse Amplitude Modulation, PAM)은 식물의 광합성 특성을 빠르게 측정하며, 민감하고 비파괴적인 기술로 널리 사용되지만, 절대적인 값의 도출이 아닌 상대적인 특성을 비교하는데 주로 사용된다(Ralph and Burchett, 1995; Maxwell and Johnson, 2000; Durako and Kunzelman, 2002). 수중에서 8–10개체의 잘피를 임의로 선정하여 두 번째 잎의 중간 부분에 클립(dark leaf clip)을 고정하였다. 광계 Ⅱ의 최대양자수율(maximum quantum yield)은 10분간 암적응(dark-adapted) 시킨 후 포화광(saturation pulse)을 조사하여 측정하였고, 다음 식 (1)을 통해 계산하였다.
이때의 F0은 암적응된 잎의 초기 형광값이며, Fm은 포화광에 의해 유도된 최대형광값이다.
빠른 광반응곡선(rapid light curves, RLCs)은 0–1,002 μmol photons m−2 s−1 범위 내에서 설정된 8단계 광조건에서, 각 단계별로 10초간 주어진 광도(actinic light)에 노출한 후 포화광을 조사하여 광계 Ⅱ의 유효양자수율(effective quantum yield, ΦPSⅡ)을 측정하고, 이 값을 이용하여 상대전자전달율(relative electron transport rate, rETR)를 계산하여 측정하였다(Ralph and Gademann, 2005). 유효양자수율(effective quantum yield, ΦPSⅡ)은 엽록소가 흡수한 빛 중에서 광화학과정에 사용한 비율을 의미하며 명적응(light-adapted)된 상태에서 측정하여 다음 식 (2)를 통해 계산하였다.
여기서 F'm은 명적응 상태에서 포화광에 의해 유도된 최대형광값이며, Ft는 명적응 후 매우 낮은 광도에서의 초기형광을 의미한다. 상대전자전달율은 다음 식 (3)으로 계산하였으며, PAR (photosynthetic active radiation, μmol photons m−2 s−1)은 8단계 각각의 광량, 0.84는 광합성 색소에 의한 광자의 흡수율, 0.5는 광자가 2개의 광계로 균등하게 흡수됨을 가정한 상수이다.
측정된 상대전자전달율 값들은 광량에 대해 도시하고 Webb et al.(1974) 또는 Platt et al.(1980)의 함수를 이용하여 피팅 후 상대적인 광합성 매개변수들[전자전달효율(α), 포화광량(Ek), 최대상대전자전달율(rETRmax)]을 계산하였다(Jassby and Platt, 1976).
2.5 통계분석
모든 측정치는 평균(mean)과 표준오차(SE)로 나타내었다. 통계분석은 SPSS 프로그램(version 20.0, SPSS, Inc., Chicago, IL, USA)을 이용하였으며, 분석 전 모든 자료는 정규성(normality)과 등분산성(homogeneity of variance)을 검정하였으며, 가정이 성립되지 않을 경우 로그로 변환하였다. 자연생육지와 이식 장소의 환경요인(수중광량, 수온, 퇴적물 내 무기영양염류) 및 잘피의 생리생태학적 특성(생육밀도, 성장률, 형태, 광합성 특성)의 조사장소 및 조사시기별 유의한 차이는 two-way ANOVA를 이용하여 분석하였으며, 해수 내 무기영양염류의 시간에 따른 유의한 차이는 one-way ANOVA를 이용하여 분석하였다(p<0.05). 분석값이 유의할 경우, Tukey 검정법을 이용하여 그 유의성을 분석하였다.
3. 결 과
3.1 환경요인
자연생육지와 이식 장소의 일일 평균 수중광량은 모두 뚜렷한 계절적 경향을 보이지 않았지만, 조사 장소에 따라 유의한 차이를 보였다(p<0.001; Fig. 2A). 2011년 10월부터 2012년 4월까지 측정된 이식 장소에서의 수중광량(7.3 mol photons m−2 d−1)은 자연생육지에서의 수중광량(3.5 mol photons m−2 d−1)보다 유의하게 더 높은 수치를 보였다(p<0.001; Fig. 2A). 두 조사 장소의 수온은 여름에 높고 겨울에 낮은 뚜렷한 온대해역의 특징을 보였지만(p<0.001; Fig. 2B), 자연생육지와 이식 장소 간의 유의한 차이는 보이지 않았다(p=0.228). 자연생육지의 수온은 조사기간 중 9.5-22.6℃로 관찰되었다.
해수 속 무기영양염 농도는 조사시기별로 유의한 차이를 보였으나(p<0.01; Fig. 3A, B, C), 뚜렷한 계절성을 나타내지 않았다. 해수 내 NH4+와 NO3−+NO2−, PO43−의 평균 농도는 각각 2.0 μM, 4.0 μM, 0.5 μM이었다. 퇴적물 공극수의 무기영양염 역시 조사시기별로 유의한 차이를 보였지만(p<0.01; Fig. 3D, E, F), 뚜렷한 계절성을 보이지 않았다. 퇴적물 공극수 내 무기영양염 농도는 자연생육지보다 이식 장소에서 유의하게 더 높은 값을 나타내었다(p<0.001). 퇴적물 공극수의 NH4+, NO3−+NO2−, PO43− 농도는 각각 자연생육지에서 4.6 μM, 6.9 μM, 0.7 μM이었으며, 이식 장소에서는 19.5 μM, 20.1 μM, 2.0 μM이었다(Fig. 3D, E, F).
3.2 왕거머리말의 생육밀도, 성장률, 형태학적 특성
왕거머리말의 생육밀도는 봄에 증가하고 가을과 겨울에 감소하는 유의한 계절변동을 보였다(p<0.001; Fig. 4). 자연생육지에서는 봄부터 초여름까지 밀도가 증가하여 7월에 최대값(115 shoots m−2)을 보였지만, 이식 장소에서는 봄에 급격히 증가하여 5월에 최대값(80 shoots m−2)을 보였다. 이후 이식 장소에서의 생육밀도는 태풍 발생 이후부터 급격히 감소하여 이식 1년 후인 2012년 9월에는 밀도가 30 shoots m−2까지 감소하였다.
자연생육지와 이식 장소에서 왕거머리말의 개체당 잎 성장률(mg DW shoot−1 d−1)은 모두 늦봄부터 초여름까지 급격히 증가하였으며, 이식 장소의 잘피가 자연생육지에 비해 유의하게 높은 잎 성장률을 보였다(p<0.001; Fig. 5A) 이식 후 2011년 10월부터 2012년 6월까지 이식 장소에서의 개체당 평균 잎 성장률은 24.8 mg DW shoot−1 d−1로 자연생육지에서의 평균 잎 성장률인 20.2 mg DW shoot−1 d−1보다 약 23% 높게 나타났다. 자연생육지와 이식 장소에서 모두 봄부터 생육밀도가 증가함에 따라 단위면적당 잎 성장률(g DW shoot−1 d−1) 또한 급격하게 증가하였으며, 개체당 잎 성장률과는 달리 두 장소 사이에 단위면적당 잎 성장률은 유의한 차이를 보이지 않았다(p=0.32; Fig. 5B).
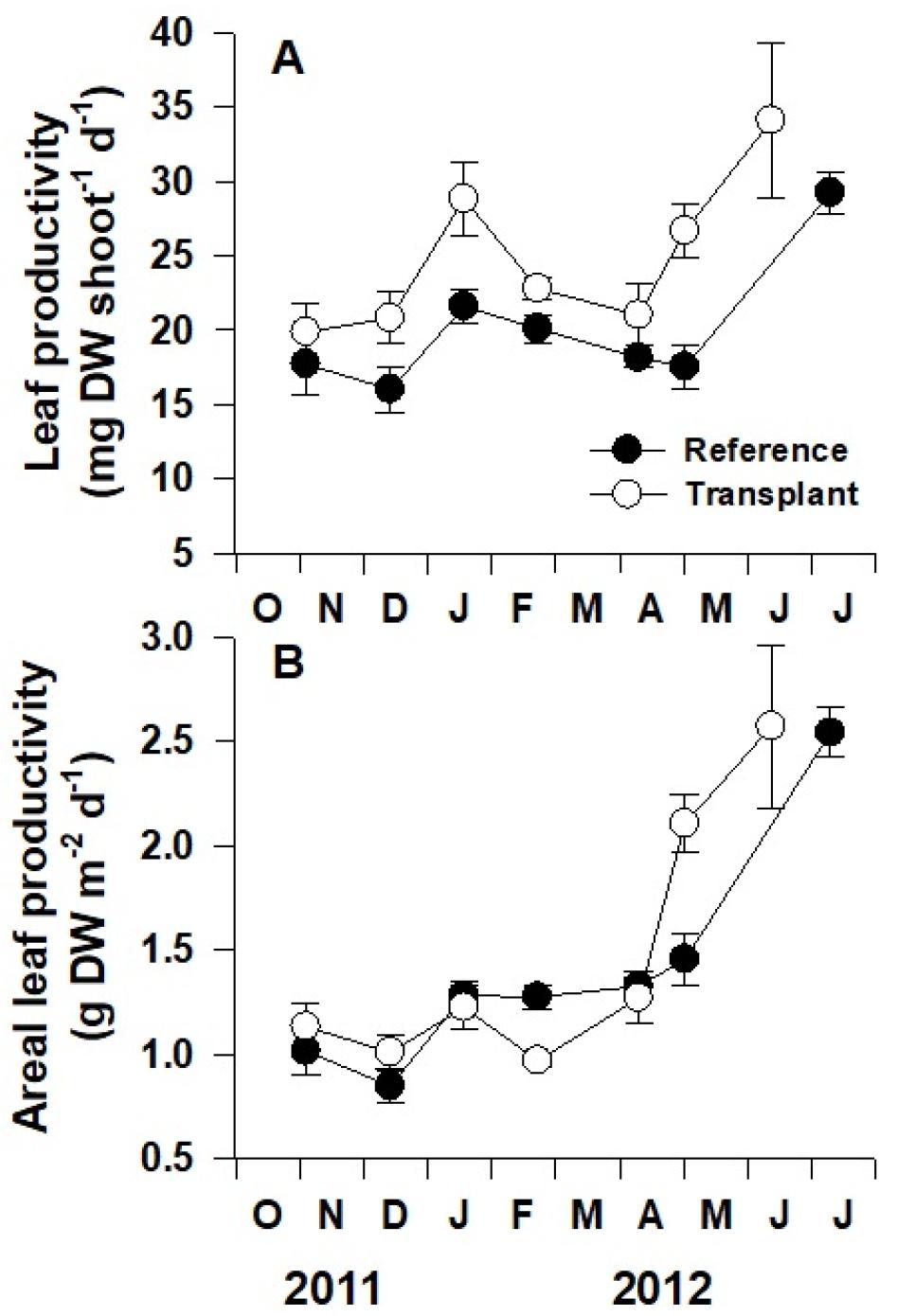
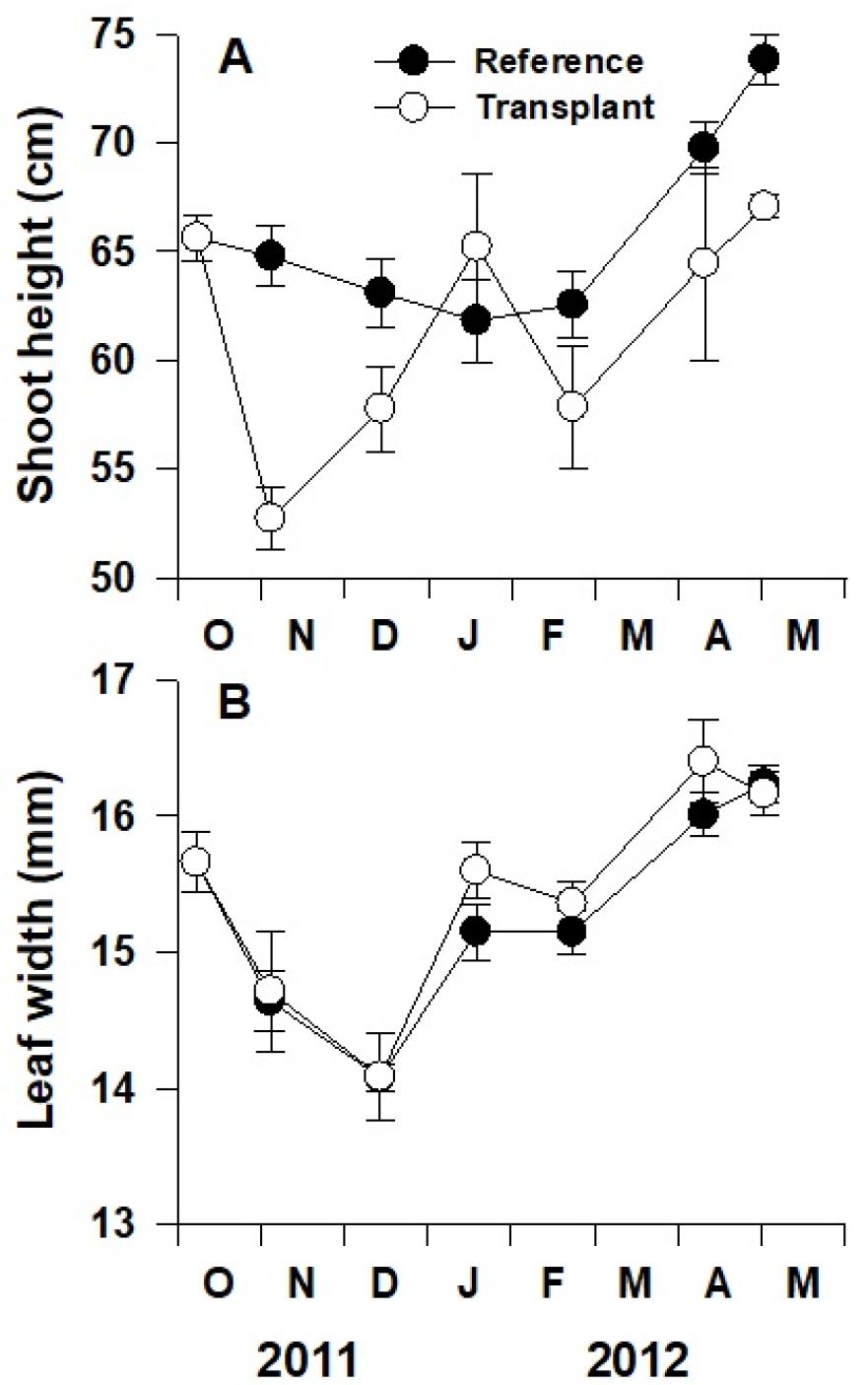
왕거머리말 잎의 전체 길이(cm)와 잎 너비(mm)는 조사시기별로 유의한 차이를 보였다(p<0.001; Fig. 6A, B). 자연생육지에서 왕거머리말의 전체 길이는 가을과 겨울에 감소하고 봄에 증가하는 뚜렷한 계절성을 보였지만, 이식 장소에서의 잘피 전체 길이는 계절 변동을 보이지 않았다. 왕거머리말의 전체 길이는 자연생육지에서 이식 장소에 비해 유의하게 높은 수치를 보였다(p<0.001; Fig. 6A). 이식된 잘피는 이식 한 달 후 현저히 작아졌다가 다시 생장하였다. 자연생육지와 이식 장소의 왕거머리말의 잎 너비는 뚜렷한 계절성을 보였으나, 두 장소 간의 유의한 차이는 관찰되지 않았다(p=0.24).
3.3 광합성 특성
최대양자수율(Fv/Fm)은 조사시기 간의 유의한 차이가 나타났으나(p<0.001), 자연생육지와 이식 장소 간의 유의한 차이는 나타나지 않았다(p=0.102; Fig. 7A). 자연생육지와 이식 장소의 왕거머리말의 평균 최대양자수율은 각각 0.814, 0.809로 조사되었다. 전자전달효율(α), 최대상대전자전달율(rETRmax), 포화광량(Ek)은 모두 조사시기(각각, p<0.01, p<0.001, p<0.001)와 장소(p<0.001)에 따라 유의한 차이를 나타냈다(Fig. 7B, C, D). 전자전달효율은 조사기간 중 뚜렷한 계절성을 보이지 않았지만, 2012년 2월을 제외하고 이식 장소보다 자연생육지보다 더 높은 수치를 보였다(Fig. 7B). 조사기간 중 평균 전자전달효율은 자연생육지에서 0.36, 이식 장소에서의 0.33의 값을 보였다. 최대상대전자전달율은 두 장소 모두 잘피 이식 실험을 시작한 2011년 10월에 가장 높은 값(15.92 μmol electrons m−2 s−1)를 보인 후 가을과 겨울에 감소하여 2012년 2월에 가장 낮은 값(자연생육지: 5.7 μmol electrons m−2 s−1, 이식 장소: 7.2 μmol electrons m−2 s−1)을 보이고 다시 봄과 여름에 증가하였다(Fig. 7C). 또한 2011년 10월과 2012년 2월, 7월에 이식 장소의 왕거머리말의 최대상대전자전달율이 자연생육지보다 유의하게 높은 수치를 보였다(p<0.001). 포화광량은 최대상대전자전달율과 마찬가지로 가을과 겨울에 감소하였다가 봄과 여름에 증가하는 계절성을 보였다. 두 장소 모두 2011년 10월에 최대값을 보였으며, 2012년 5월에 최소값을 보였다(자연생육지: 19.7 μmol photons m−2 s−1; 이식 장소: 23.3 μmol photons m−2 s−1). 조사기간 중 이식 장소의 왕거머리말 포화광도가 자연생육지보다 유의하게 더 높은 수치를 보였다(p<0.001).
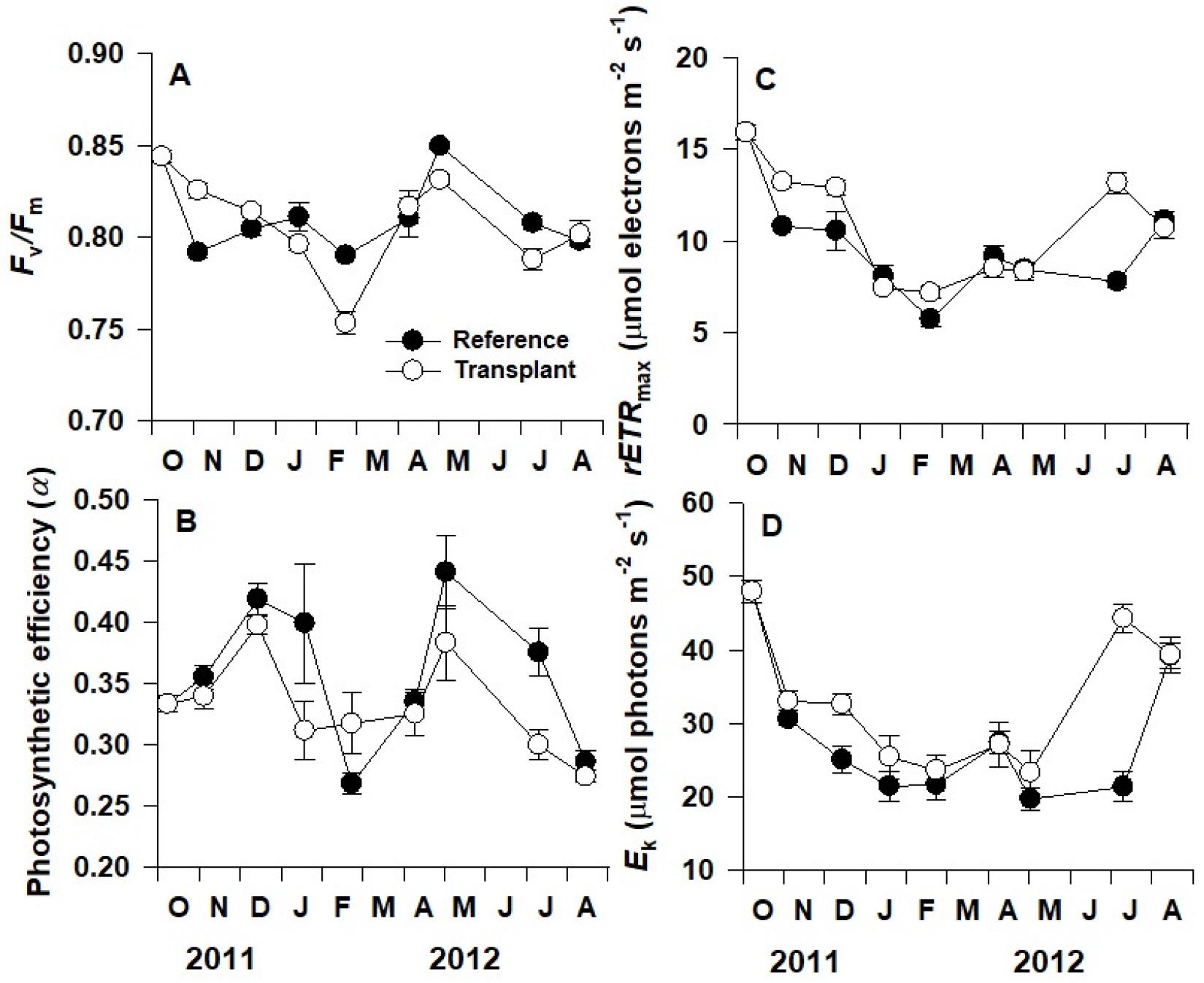
Fig. 7.
Changes in maximum quantum yield (Fv/Fm; A), photosynthetic efficiency (α; B), maximum relative electron transport rate (rETRmax,; C), and minimum saturating irradiance (Ek; D) of Zostera asiatica in the reference and transplant sites on the eastern coast of Korea from October 2011 to August 2012. Values are mean ± SE.
4. 고 찰
왕거머리말이 동해 연안의 깊은 수심에 한정되어 분포하는 이유를 밝히기 위하여, 얕은 수심에 이식한 후 이식개체들의 생장을 자연생육지의 개체들과 비교 분석하였다. 이식된 잘피는 새로운 장소에 이식하는 과정에서 잘피가 공기 중으로 노출되거나 조직 일부가 손상을 받는 등의 스트레스를 받게 된다. 이러한 환경 스트레스는 이식 후 성장률과 형태 뿐만 아니라 생존율에도 영향을 줄 수 있다(Park et al., 2009; Li et al., 2014). Park et al.(2009) 및 Li et al.(2014)의 연구에 따르면, 거머리말의 이식 후 이식 스트레스에 의해 생육밀도 및 전체 길이가 감소한다고 보고되었다. 본 연구에서도 2011년 10월 왕거머리말의 이식 후 한 달 동안 이식개체의 길이가 급격하게 감소하였다. 이는 같은 시기의 자연개체의 길이가 1.3% 감소한 것보다 더 큰 폭의 감소(19.6%)를 보여 이식 스트레스를 받은 결과로 추정되며, 이후 길이가 점점 증가하여 자연생육지의 개체들과 유사하게 되었다. 자연생육지 왕거머리말의 생육밀도는 가을에 감소하였고, 이후 겨울부터 점점 증가하는 경향을 보였지만, 이식 장소의 경우 겨울까지 지속적으로 감소하였다. 이식된 왕거머리말 생육밀도는 봄에 자연생육지와 유의한 차이를 보이지 않았지만, 이후 자연생육지에 비해 급격히 감소하였다.
잘피는 광조건이 변화하면 적응하기 위해 다양한 생리적 변화를 나타낸다(Collier et al., 2008; York et al., 2013). 해조류 및 잘피는 강한 빛에 노출되었을 때 광손상이 일어나며, 광계Ⅱ의 광보호 작용으로 최대양자수율이 감소한다(Ralph and Burchett, 1995; Durako and Kunzelman, 2002; Phandee and Buapet, 2018). 잘피는 수중광량이 증가할수록 전자전달율과 포화광도는 증가하지만, 전자전달효율은 감소한다고 보고되었다(Campbell et al., 2007; Sharon et al., 2011; Park et al., 2016b). Sharon et al.(2011)의 연구에 따르면 홍해 해역 얕은 수심의 Halophila stipulacea 종이 깊은 수심보다 더 높은 상대전자전달율과 포화광도 값을 보였다. 또한 조간대에 분포하는 거머리말은 조하대에 비해 전자전달효율은 낮지만, 상대전자전달율 및 포화광도는 높았다(Park et al., 2016b). 본 연구에서는 얕은 수심으로 이식한 잘피의 최대양자수율이 자연개체와 유의한 차이를 보이지 않아 수중광량의 증가로 인한 광손상은 일어나지 않은 것으로 추정된다. 또한 이식된 왕거머리말은 자연생육지 개체들에 비해 전자전달효율은 낮았지만, 최대상대전자전달율 및 포화광도는 오히려 증가하였고, 이는 얕은 수심에서 수중광량의 증가로 인한 잘피의 생리적 변화로 판단된다.
얕은 수심으로 이식된 왕거머리말은 이식초기에 크기 및 생육밀도가 자연생육지 개체에 비해 감소함에도 불구하고, 개체당 일일 성장률은 오히려 증가하였다. 잘피의 성장률은 주로 수중광량, 수온 및 무기영양염 농도에 의해 조절된다. 잘피의 광합성, 생장 및 분포 수심은 수중광량과 직접적으로 연관되어 있으며, 수중광량이 증가할수록 잘피의 성장률이 증가한다고 보고되었다(Lee et al., 2007b; O’Brien et al., 2018). 이식 장소는 자연생육지보다 수심이 얕아 조사기간 중 평균 수중광량이 2배 이상 높았다(이식 장소: 7.3 mol photons m−2 d−1; 자연생육지: 3.5 mol photons m−2 d−1). 이전 연구에서 동해안 왕거머리말의 성장률은 수중광량과 유의한 연관이 있으며, 수중광량이 증가할수록 성장률이 증가한다고 보고된 바 있다(Park et al., 2016a). 또한 퇴적물 공극수 내 무기영양염 농도 역시 이식 장소가 자연생육지보다 유의하게 높은 수치를 보였으며 평균 NH4+ 농도는 약 4.2배, NO3−+NO2−와 PO43−는 각각 2.9배, 2.7배 높았다. 대부분의 잘피생육지는 낮은 무기영양염 농도로 인해 생장이 제한되며, 무기영양염 농도가 증가하면 잘피의 생장이 증가한다고 알려져 있다(Short, 1987; Lee et al., 2007b). 따라서, 조사해역 얕은 수심의 잘피생육지 환경조건은 수중광량 및 무기영양염 농도 증가로 인해 오히려 생장에 더 유리하다고 판단된다.
본 연구에서 이식된 왕거머리말은 새로운 환경에 노출되어 다양한 환경에 적응하기 위해 다양한 생리적·생태학적 변화가 나타났다. 얕은 수심에 이식된 개체들의 광합성 특성, 성장률 및 형태적 특성들이 자연 개체들과 유사하게 유지되었으나 2012년 8-9월에 태풍 덴빈(최고풍속 162 km/h, 중심기압: 945 hPa)과 산바(최고 풍속: 202 km/h, 중심기압: 910 hPa)의 영향으로 이식 장소의 개체 대부분이 소실되었다. 국립해양조사원 왕돌초 조위관측소(울진군 후포항 동쪽 24.9 km)의 2012년 기록에 의하면 파주기 7초 이상의 너울성 파도가 월평균 9일에서 31일 동안 발생하였다. 우리나라 동해안은 해안선이 단조로워 내만이나 폐쇄된 해역이 거의 없어, 얕은 수심에서는 태풍이나 너울성 파도에 의한 물리적 스트레스를 받기 쉽다. 따라서 얕은 수심에서의 파랑에너지로 인해 잘피생육지가 깊은 수심에 제한적으로 형성될 수 있다(Infantes et al., 2009; Vacchi et al., 2010). Vacchi et al.(2010)에서는 파도가 부서지는 얕은 수심에서 잘피생육지가 분포하지 못한다고 보고하였다. 이 연구에 따르면 파도가 부서지는 수심(쇄파수심; breaking depth) 이하에서는 죽은 잘피들만 간혹 발견되었으며, 파도가 부서지지 않는 깊은 수심(closure depth)에서 Posidonia속 잘피생육지가 잘 형성되었다. 우리나라 동해안의 왕거머리말은 개방된 연안에서 8 m 이상의 깊은 수심에 분포하지만, 일본 아케시만의 왕거머리말은 비교적 얕은 수심(1.5-3.0 m)에 분포하고 있으며, 이는 파랑에너지로부터 보호받을 수 있는 내만에 위치하고 있기 때문이라고 볼 수 있다(Lee et al., 1999). 왕거머리말 잎의 폭은 다른 거머리말 속의 잘피종들에 비해 약 1.4-1.6배 넓으며, 이는 깊은 수심에서의 낮은 광조건에서 빛을 효율적으로 흡수하기 위한 적응의 결과로 판단된다. 잘피들은 다양한 형태의 치명적인 자연재해(catastrophic events)에 의해 그 분포가 한정될 수 있다. 갑작스럽게 발생하는 태풍, 지진, 쓰나미, 홍수 및 수온 변화(marine heat wave; La Niña), 동물들의 과도한 섭식활동(overgrazing), 조류 번성(algal bloom)으로 인한 수중 광량의 감소 등으로 인해 대규모 잘피생육지 감소가 보고되었다(Peterson et al., 2002; Campbell and McKenzie, 2004; Lee et al., 2007a; Rivers and Short, 2007; Kim et al., 2015; Thomson et al., 2015; Moseby et al., 2020). 본 연구 결과에 의하면 얕은 수심에 이식된 왕거머리말이 평상적인 환경 조건하에서는 비교적 잘 생육하였으나, 태풍과 같은 자연재해 발생시 강한 물리적 에너지에 의해 개체들이 뽑혀 나가 얕은 수심에서는 분포가 불가능한 것으로 추정되었다. 파랑에 의한 물 입자의 운동은 수심이 깊어질수록 감소하며, 일정한 수심 이하에서는 사라지게 되어 그 영향을 적게 받을 수 있다(Koch et al., 2007). 따라서 우리 동해 연안의 왕거머리말은 태풍 등과 같은 비정상적으로 강한 파랑에너지 발생시 그 영향을 피하기 위하여 깊은 수심에 한정되어 분포하고 있는 것으로 판단되었다.
