

1. 서 론
2. 재료 및 방법
2.1 종자 준비
2.2 건조(노출)시간에 따른 종자의 발아율과 함수율
2.3 온도에 대한 종자의 발아율
2.4 통 계
3. 결 과
3.1 건조(노출)시간에 따른 거머리말 종자의 발아율과 함수율
3.2 온도에 대한 종자의 발아율
4. 토 의
1. 서 론
거머리말(Zostera marina L.)은 북반구에서 가장 넓게 분포하는 잘피(seagrasses)이다. 또한, 우리나라 연안의 우점하는 종으로 잘피로 통칭되기도 하며, 특히 남해안에 넓은 군락지를 형성하고 있는 해양성 침수식물이다(den Hartog, 1970; Lee and Lee, 2003). 우리나라 연안에서 거머리말은 조간대부터 수심 5 m까지 조하대에 나타나며, 남해안에서는 해안선을 따라 대부분 2-3 m의 낮은 수심에 출현한다(Lee and Lee, 2003). 거머리말은 일차생산자로 광합성을 통하여 높은 생산성을 발휘하며 지구상의 가장 생산성이 높은 생태계 중 하나로 연안생산성에 기여하고 있다(Duarte, 2000). 거머리말 서식지는 다양한 생물들의 산란장과 서식지를 제공하여 수산생산성을 향상시키고, 해수 중의 영양염을 제거하여 해양환경을 정화시키는 기능도 수행한다(Hovel et al., 2002; Thomas and Cornelisen, 2003). 그러나 최근 우리나라뿐만 아니라 세계적으로도 많은 거머리말 서식지가 인위적인 요인으로 훼손되고 있다(Fonseca et al. 1998; Park and Lee, 2007). 거머리말 서식지를 위협하는 주요 요인으로는 매립, 해안선 공사, 육상으로부터 과도한 영양염의 공급 등이며, 비교적 얕은 수심에 넓은 면적으로 분포하는 거머리말은 서식지 훼손이 가장 빈번한 잘피종이다(Orth et al., 2006; Shafer and Bergstrom, 2010). 또한 거머리말은 국내외에서 서식지 복원이 가장 많이 시도된 잘피종으로, 다양한 거머리말 성체 이식방법이 개발되어 서식지 복원사업에 활용되어 왔다(Park and Lee, 2007). 그러나 성체를 이식하는 방법은 공급용 거머리말 군락지에 일시적인 훼손이 불가피하고, 잠수작업이 필수적으로 포함되는 경우가 많아 서식지 복원을 위해 고비용이 소요되는 경우가 빈번하였다(Fonseca et al., 1998). 이에 비해 종자를 파종하여 잘피서식지를 복원하는 방법은 기존 잘피장의 훼손을 줄일 수 있고 단기간에 넓은 서식지를 복원할 수 있으며, 복원 비용이 저렴하다(Tanner and Parham, 2010). 이러한 장점들로 말미암아 2000년대 이후부터 종자를 활용한 거머리말 서식지 복원 사업이 체스픽만 등에서 본격적으로 시도되고 있다(Shafer and Bergstrom, 2010).
종자를 활용한 거머리말 서식지 복원을 위해서는 발아 가능한 건강한 거머리말 종자의 확보와 관리가 우선되어야 한다. 침수식물인 거머리말은 대부분 해수 중에서 생활하며, 공기에 노출 시 건조 스트레스를 받게 된다. 거머리말 성체가 받는 건조 스트레스의 형태로 광합성 효율이 저하되거나 잎의 괴사가 밝혀졌다(Shafer et al., 2007). 조하대에 서식하는 거머리말에 비해 일정기간 공기중 노출되는 조간대에 서식하는 거머리말은 잎의 노출로 인한 건조 스트레스로 개체의 형태와 생체량이 현저히 줄어드는 것으로 보고되고 있다(Kim et al., 2013). 그리고 거머리말은 해수 중에서 개화, 수분, 열매의 형성과 종자 성숙이 진행되며 함수율이 높은 종자를 생산한다. 거머리말 성체에서 건조 스트레스가 보고되는 것처럼 거머리말 종자도 공기 중 노출로 인한 종자의 건조가 나타날 수 있으며, 이는 발아율의 감소나 종자의 사망으로 이어질 수 있을 것이다. 최근 거머리말 종자를 건조시킨 후 저염의 해수에서 발아한 자료가 보고되고 있으나(Pan et al., 2012), 아직도 건조에 의한 거머리말 종자의 발아율이나 함수율에 대한 자료는 매우 부족하다.
종자를 활용한 거머리말 서식지 복원 과정에서 거머리말 종자는 채취, 보관과 현장 파종을 위한 준비 작업등을 거치게 되며, 이들 작업 과정 중 일정 기간 공기 중 노출될 수 있다. 노출 중 한계 범위 이상의 건조 스트레스를 겪어 활력이 감소한 거머리말 종자는 서식지 복원의 목적으로 부적합할 수 있을 것이다. 본 연구에서는 거머리말 종자의 한계 함수율을 알아보기 위하여, 종자의 건조로 인한 함수율과 발아율의 변화를 측정하였다. 이 결과는 서식지 복원을 위한 거머리말 종자의 취급과 관리에 중요한 정보를 제공하고자 한다.
2. 재료 및 방법
2.1 종자 준비
거머리말의 화지는 남해안의 진동만(35° 06´N, 128° 32´E)과 거제만(34° 48´N, 128° 35´E)에서 2015년 6월말부터 7월초에 SCUBA를 이용한 잠수작업으로 채취하였다. 채취된 화지들은 현장 해수가 담긴 아이스박스에 넣어 실험실로 운반하여 해수가 지속적으로 투과되는 망에 넣어 보관하였고, 약 1개월 후 화지조각과 이물질을 제거하고 종자를 수집하였다. 수집한 종자는 실온에서 공기가 지속적으로 공급되는 해수(32 psu)에 보관하였으며, 실험에 앞서 채칩된 거머리말 종자의 활력을 알아보기 위해 테트라졸리움 검사(Tetrazolium test)를 실시하였다(Kauth and Biber, 2014). 즉, 거머리말 종자의 종피를 제거하고 0.08% 테트라졸리움에 24시간 침지 후 종자의 단면을 잘라 염색여부를 확인하였으며, 배 부분이 붉게 염색된 것을 활력이 있는 것으로 간주하였다(n=100).
2.2 건조(노출)시간에 따른 종자의 발아율과 함수율
공기 노출에 따른 건조된 거머리말 종자의 발아율을 조사하기 위해 거머리말 종자(n=100)를 상대습도 50%, 온도 20°C의 항온 항습기에 넣은 후 실험 설계된 시간 단위로 건조 시킨 후 발아율을 측정하였다(n=3). 거머리말 종자는 0.5시간부터 5시간까지 30분 단위로, 이후 6시간부터 15시간까지 1시간 단위로 공기에 노출시켰다. 조건별로 노출시킨 후 거머리말 종자는 즉시 여과해수(32 psu, GF/C, Watman, Maidstone, England) 50 mL를 주입한 페트리디쉬(직경 9 cm)에 넣은 후 실온(17°C)에서 45 µmol m-2s-1 (12L:12D) 광조건 하에 발아실험을 실시하였다(Park et al., 2012). 거머리말은 종피(seed coat)가 벗겨져 떡잎(cotyledon)이 나타난 것을 발아한 것으로 간주하였다(Park et al., 2012).
서식지 복원을 위한 종자의 채취 및 취급 과정에 필요한 종자 관리 정보를 얻기 위해 공기 노출에 따른 건조된 거머리말 종자의 함수율을 조사하기 위해 거머리말 종자 5개씩을 상대습도 50%, 온도 20°C의 항온항습기에 0.5시간부터 5시간까지 30분 단위로, 이후 6시간부터 15시간까지 1시간 단위로 공기에 노출시켰다. 각 노출시간 직후 거머리말 종자의 습중량을 측정한 뒤 건조기(60°C)에서 48시간 건조 후 건중량을 측정하여, 거머리말 종자의 함수율을 구하였다.
2.3 온도에 대한 종자의 발아율
온도에 대한 거머리말 종자의 발아율을 측정하기 위해 거머리말 종자(n=100)를 상대습도 50%로 고정된 10°C, 20°C 및 30°C의 항온항습기에 1, 2, 4, 8, 16, 24시간 건조시켰다(n=3). 각 온도의 항온항습기에 건조된 거머리말 종자는 각 노출시간 후 즉시 발아실험을 실시하였다. 각 조건에서 건조된 거머리말 종자의 발아율 측정은 상기의 상대습도 50%, 온도 20°C의 공기에 노출된 거머리말 종자의 발아율 실험과 동일한 방법으로 수행하였다.
2.4 통 계
모든 측정치는 평균(mean)과 표준오차(SE)로 나타내었다. 모든 자료는 normality와 homogeneity of variance를 검정한 후 노출 시간에 따른 거머리말 종자의 함수율과 발아율 및 다양한 온도에 노출된 거머리말 종자의 발아율을 Two-way ANOVA를 이용하여 분석하였다. 노출시간에 따른 거머리말 종자의 함수율과 발아율의 상관관계는 Pearson’s correlations를 이용하여 분석하였다. 분석값이 유의할 경우, Turkey HSD (Honestly Significant Difference)를 이용하여 각 자료의 유의성을 검증하였으며, 통계분석은 SAS 9.1을 이용하였다.
3. 결 과
3.1 건조(노출)시간에 따른 거머리말 종자의 발아율과 함수율
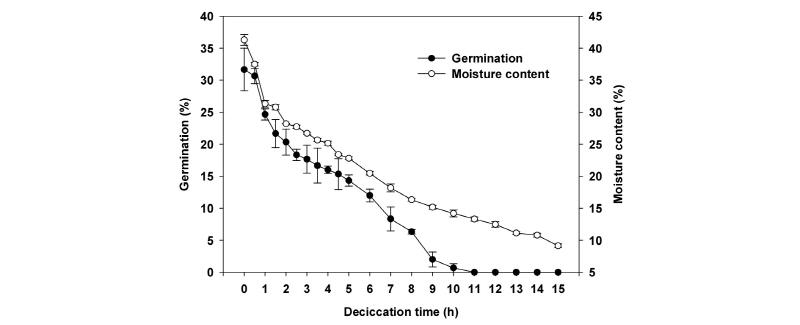
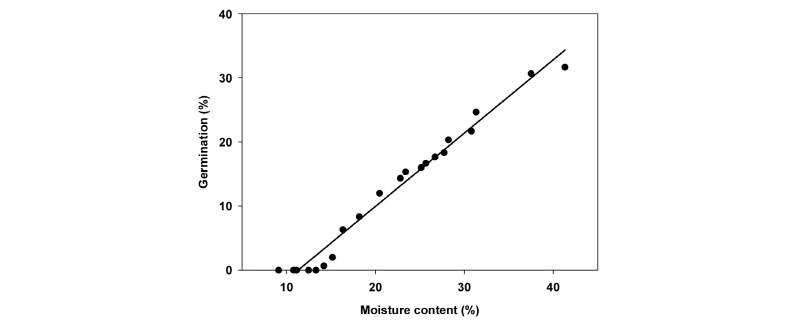
테트라졸리움 시험 결과 거머리말 종자는 모든 종자의 배 부분이 붉게 염색되어 100%의 활력을 보였다. 신선한 거머리말 종자의 습중량은 11.44±0.48 mg으로, 31.7±3.3%의 발아율과 41.3±0.8%의 함수율을 나타내었다. 종자가 공기에 노출되는 시간이 길어질수록 거머리말 종자의 발아율과 함수율은 급격한 감소를 보이는 음의 상관관계가 나타났고(발아율: r2=0.9088, F=189.30, p<0.001; 함수율: r2=0.9127, F=198.71, p<0.001), 함수율(y=-33.52-5.48x)에 비해 발아율(y=-25.53-6.33x)이 더욱 급격히 감소하였다(Fig. 1). 거머리말 종자의 발아율은 공기 노출 1시간 이후(종자 함수율 31.33±0.52%)부터 감소되었으며, 11시간 이후(종자 함수율 13.31±0.33%)에는 전혀 발아하지 않았다. 그러나 건조된 종자의 함수율과 발아율 간에는 양의 상관관계(r2=0.9792, F=894.49, p<0.001)가 나타나 종자의 함수율이 높을수록 발아율도 증가(y=-12.94+1.14x; y=발아율, x=함수율)하였다(Fig. 2).
|
Fig. 1. Changes of germination rate and moisture content of Z. marina seeds exposed at relative humidity of 50% under 20°C. |
|
Fig. 2. Linear relationship between moisture content and germination rate of Z. marina seeds exposed at relative humidity of 50% under 20°C. |
3.2 온도에 대한 종자의 발아율
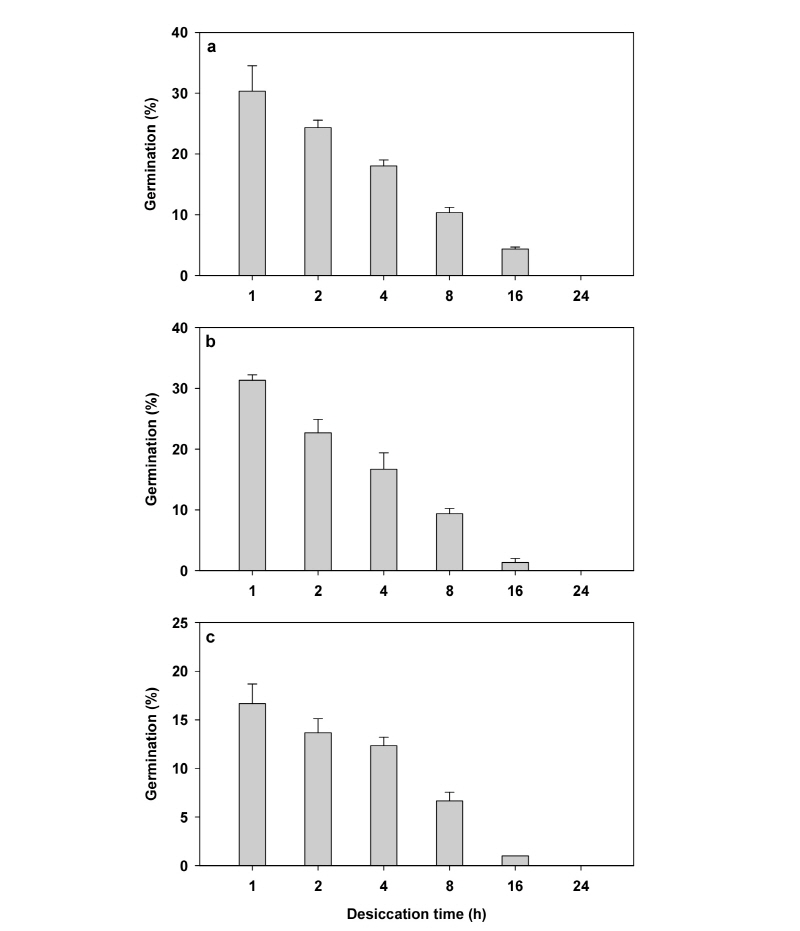
온도는 거머리말 종자의 발아율에 유의한 영향을 미쳤다. 모든 온도에서 노출시간이 길수록 거머리말 종자의 발아율은 현저히 감소하였으며, 24시간 공기에 노출된 종자는 전혀 발아하지 않았다. 24시간을 제외한 모든 노출 시간에서 10°C와 20°C에 노출된 거머리말 종자의 발아율이 30°C에 노출된 거머리말 종자의 발아율보다 유의하게(p<0.001) 높았다(Fig. 3).
4. 토 의
실험 결과 공기에 노출된 거머리말 종자는 함수율과 발아율의 감소가 나타났다. 건강한 거머리말 종자는 41.3%의 평균 함수율을 보유하여, 일반적으로 10% 미만의 함수율을 지니는 건조 종자(orthodox seeds)보다는 30-70%의 함수율을 지니는 난저장성 종자(recalcitrant seeds)에 해당되었다(Chin, 1995). 건조 종자는 공기에 노출되어도 종자 활력이 거의 상실되지 않는데 비해 난저장성 종자는 건조에 민감하여 한계 함수율 이하로 일정시간 노출되면 종자내 조직이 변형 또는 파괴되어 종자 활력이 상실된다(Hay et al., 2000). 난저장성 종자의 한계 함수율은 식물 종에 따라 다르게 나타나고, 동종의 식물에서도 서식환경에 따라 다르게 나타날 수 있다(Kauth and Biber, 2014). 본 결과에서 건강한 거머리말 종자의 발아율은 31.7%, 공기 노출 30분 후는 30.7%로 유사하였으나, 노출시간이 1시간 이후부터 발아율이 감소(24.7%)하였으며, 11시간 이상 노출된 거머리말 종자는 전혀 발아하지 않았다. 즉, 공기 노출 1시간의 거머리말 종자(함수율 31.33%)는 건강한 거머리말 종자(함수율 41.30%)와 30분 노출된 거머리말 종자(37.5%)에 비해 각각 24.1%와 16.5%의 함수율 감소가 나타났으며, 이때 발아율도 각각 22.1%와 19.7%가 감소하였다. 따라서 본 실험 결과 거머리말 종자의 발아 한계 함수율은 31.3% 이상 37.5%미만의 범위로 추측할 수 있다. Pan 등(2012)도 상대습도 50%, 온도 25°C에 2시간 노출된 거머리말 종자는 노출되지 않은 종자에 비해 활력이 51.1%, 함수율이 33.5% 감소한 것으로 보고하고 있다. 다만, 거머리말 종자의 정확한 한계 함수율의 파악을 위해서는 1시간 이내의 노출시간에서 더 세분화된 시간 단위로 건조된 종자의 발아율 관찰이 요구된다.
본 실험에서 10°C, 20°C 및 30°C에 노출된 거머리말 종자의 발아율은 유의한 차이를 나타내었다. 10°C와 20°C에 노출된 거머리말 종자에 비해 30°C에 노출된 종자의 발아율은 24시간을 제외한 모든 노출시간에서 현저히 낮은 값을 나타내었다. 이는 거머리말 종자의 노출 온도가 종자의 발아율에 영향을 주며, 거머리말 종자는 고온에서 더 높은 건조 스트레스를 받는 것으로 추측할 수 있다. 공기에 노출되지 않은 건강한 거머리말 종자의 발아율도 저온에서 높은데 비해 고온에서 낮게 보고되었다(Park et al., 2012). 따라서, 건강한 거머리말 종자의 발아율뿐만 아니라, 거머리말 종자가 노출된 온도도 거머리말 종자의 발아율에 영향을 준다고 할 수 있다. Pan 등(2012)도 15°C부터 35°C까지 5°C간격으로 거머리말 종자를 건조시킨 후 저염(5 psu)의 해수에 발아시킨 결과 저온에서 건조한 종자의 발아율이 고온에서 건조한 종자보다 높아(Table 1) 본 결과와 유사하게 나타났다.
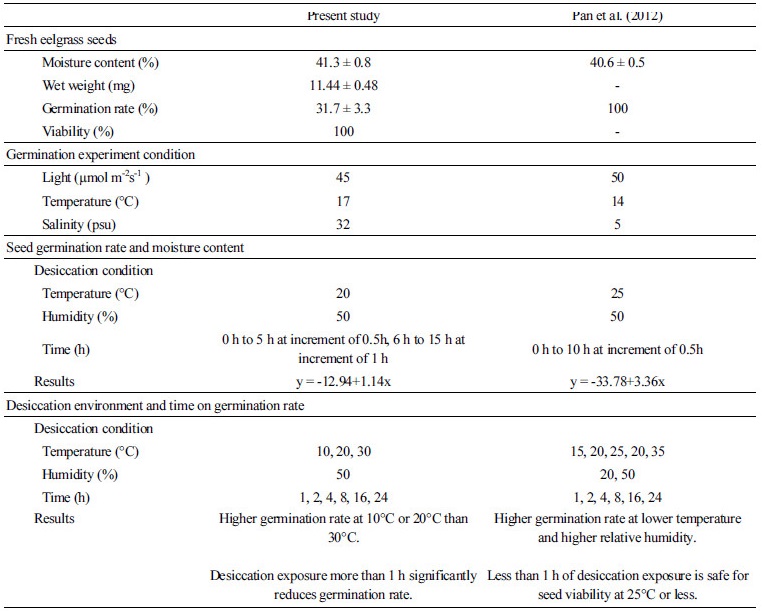
거머리말 종자는 염분, 수온, 빛 등의 환경 조건에 따라 발아율의 차이가 크게 나타나는 것으로 알려져 있다(Churchill, 1983). 본 연구는 17°C의 온도와 32 psu의 해수에서 거머리말 종자의 최대 평균 발아율은 약 31.7±3.3%로, 이는 5°C와 0 psu의 해수에서의 발아율 82.8±2.9%(Park et al., 2012), 14°C와 5 psu의 해수에서의 발아율 83.3±3.5%(Pan et al., 2011), 15°C와 5 psu의 해수에서의 발아율 100%(Pan et al., 2012)보다 낮게 나타났다. 저염 또는 무염 환경은 잘피 종자 조직의 수화를 촉진시켜 발아율을 향상시키는 데 효과적(Park et al., 2012)일 수는 있으나, 저염 또는 무염에서 발아한 거머리말 종자가 유식물로 성장하기 전 곰팡이류 등의 번식으로 사망하는 결과가 관찰되었다(Personal observations, 2015). 본 연구는 저염 해수가 아닌 일반 염분 해수에서 발아 실험이 이루어져 최대 평균 발아율이 비교적 낮게 나타난 것으로 판단된다.
난저장성 종자를 생산하는 대부분의 수생식물은 식물종과 서식환경에 따라 성체에서도 건조 내성의 정도가 다양하게 나타난다(Kauth and Biber, 2014). 침수식물인 잘피도 종에 따라 건조내성 정도가 다르며, 공기 중 노출된 다양한 종의 잘피 성체는 40-80%의 함수율 감소를 보였다(Tanaka and Nakaoka, 2004). 거머리말 성체의 경우 상온의 공기에 15-20분 노출되면 약 65%의 수분이 감소되며 이때 광합성을 멈춘다고 한다(Leuschner et al., 1998). 본 실험에서도 20°C의 공기에 1시간 이상 노출된 거머리말 종자는 발아율이 현저히 감소하기 시작하였다. 따라서, 거머리말 성체에서 건조 스트레스가 밝혀진 것처럼 거머리말 종자도 유사한 건조 스트레스가 나타남을 확인할 수 있었다. 최근 종자를 활용한 거머리말 서식지 복원 사업이 다양하게 시도되고 있는데 본 실험의 결과는 이러한 거머리말 서식지 복원 사업에 종자 취급의 기초자료를 제공할 것으로 기대된다.
